Розділ 7.експериментальні дослідження впливу різних форм металів на перебіг вагітності, плаценто-та ембріогенез

Показники загальної ембріональної смертності в контрольній групі тварин становили 11,11±4,43%, що знаходиться в межах спонтанного рівня, притаманного білим щурам та відповідає результатам досліджень інших авторів, так само, як і показники дота постімплантаційної смертності — 0,10±0,05 од. і 0,01±0,01 од. відповідно. Введення ацетату свинцю протягом усього терміну вагітності призвело до збільшення… Читати ще >
Розділ 7.експериментальні дослідження впливу різних форм металів на перебіг вагітності, плаценто-та ембріогенез (реферат, курсова, диплом, контрольна)
Нашими попередніми еколого-гігієнічними, епідеміологічними та натурними клініко-гігієнічними дослідженнями встановлено екологічну детермінованість репродуктивного здоров’я мешканців промислового регіону. Підвищене техногенне навантаження ксенобіотиками організму людини зумовлює виражені зміни його мікроелементного статусу, що, в свою чергу, супроводжується розвитком ускладнень під час вагітності і пологів.
Для розуміння патоморфологічних механізмів розвитку вищезазначених репродуктивних ускладнень нами проведено серію експериментальних досліджень із застосуванням провідних на усіх ланках міграційних процесів асоціацій ВМ — свинець-цинк у їх органічних і неорганічних формах. Слід зазначити, що така комбінація металів обґрунтована широким їх розповсюдженням у об'єктах довкілля промислового регіону за взаємопротилежних тенденцій — при підвищеному рівні надходження свинцю до організму жінок промислового регіону спостерігається виражений дефіцит цинку в харчуванні та внутрішньому середовищі організму [15, 23, 24, 107, 155, 233].
Аналіз існуючих на сьогоднішній день експериментальних робіт щодо вивчення ембріотоксичної дії свинцю здебільш стосуються високих його концентрацій — на рівні чи дещо нижче LD50, які практично не зустрічаються в реальних умовах. У той же час експериментальних робіт, присвячених вивченню впливу низьких концентрацій свинцю на ембріональний розвиток вкрай мало [119, 129, 131]. Питання морфології та патогенезу такого впливу на плаценту і плід залишаються практично невивченими, особливо у порівняльному аспекті дії неорганічних та органічних форм свинцю, а також у комбінації з природним його біоантагоністом — цинком, що і зумовлило актуальність даних досліджень.
Вплив низьких доз важких металів на організм самиці щура Вплив досліджуваних ВМ на організм самиці щура оцінювали за комплексом фізіологічних, макроскопічних, морфологічних, морфометричних, гістоморфометричних і фізико-хімічних досліджень, що дозволило встановити патоморфологічні особливості їх ізольованої та комбінованої дії на перебіг вагітності і плацентогенез, а також виявити залежність від хімічної форми металу.
У результаті проведених досліджень встановлено, що загальний стан самиць щурів усіх дослідних груп та їх поведінка суттєво не змінювались під час есперименту. Тварини були активними, вставали на задні лапки, принюхувались, умивались, з цікавістю реагували на зовнішні подразники, відносно спокійно, без агресії, сприймали процедуру введення внутрішньошлункового зонду, активно споживали їжу та воду.
Подібна ситуація характерна для іншого інтегрального показника загального стану організму — ректальної температури (табл. 7.1), яка у тварин усіх експериментальних груп за середніми значеннями коливалась у фізіологічних межах — 37,2±0,17−38,0±0,27 °С [213]. Проте лише у тварин контрольної групи протягом усього терміну вагітності спостерігалося статистично значуще зниження ректальної температури на 0,61 °С — з 37,85±0,25 °С до 37,24±0,16 °С, що фізіологічно детерміновано змінами гормонального фону.
У дослідній групі № 4 даний показник наприкінці вагітності виявився на 0,75 °С вищим (р<0,05) порівняно з контрольною групою та групою № 1, що може свідчити про певні пристосувально-компенсаторні метаболічні зміни в організмі самиці під час вагітності. Слід відмітити динаміку ректальної температури в дослідній групі, яка отримувала цитрат цинку. Вона характеризувалась тенденцією до зниження наприкінці 1-го тижня вагітності з послідуючим підвищенням на 0,74 °С (р<0,05) на 2-му тижні вагітності зі стабілізацією показника наприкінці вагітності. Порівнюючи отримані дані з динамікою маси тіла вагітних самиць даної дослідної групи за аналогічний період (табл. 7.2), можна помітити певну суперечність — при максимальному прирості маси за 1-й тиждень вагітності серед усіх досліджуваних груп — 14,63±4,6 г спостерігаються найнижчі показники ректальної температури. Така ситуація, на нашу думку, має цілком логічне пояснення — під впливом цитрату цинку відбувається певна стимуляція приросту маси тіла на початковому періоді, що характеризується більшими енерговитратами, а відтак призводить до зниження температури тіла.
Таблиця 7.1.
Динаміка ректальної температури тіла вагітних самиць щурів (M±m).
№ групи. | Ректальна температура, °С. | ||||
початкова. | термін вагітності, дні. | ||||
1 (Pbа). | 37,3±0,17. | 37,3±0,16. | 37,33±0,19. | 37,25±0,24. | |
2 (Znх). | 37,2±0,24. | 37,5±0,29. | 37,45±0,32. | 37,48±0,16. | |
3 (Znц). | 37,23±0,31. | 36,54±0,29. | 37,28±0,25**. | 37,26±0,2. | |
4 (Pbа+Znх). | 37,5±0,28. | 37,3±0,41. | 37,36±0,31. | 38,0±0,27***. | |
5 (Pbа+Znц). | 37,2±0,17. | 37,35±0,2. | 37,23±0,16. | 37,33±0,20. | |
6 (К). | 37,85±0,25. | 37,69±0,27. | 37,33±0,28. | 37,24±0,16*. | |
7 (Pbц). | 37,89±0,27. | 37,13±0,23. | 37,05±0,24. | 37,38±0,34. | |
Примітки: * - р<0,05 порівняно з початковим значенням; ** - р<0,05 порівняно з першим тижнем вагітності; *** - р<0,05 порівняно з контрольною та дослідною групою № 1.
Морфометричні показники маси тіла інтактних самиць за 20 днів експерименту характеризуються поступовим збільшенням на 59,63±6,03 г, або 32,19% (табл. 7.2), що відповідає фізіологічним нормам для цієї вікової групи [111]. Отримані нами дані динаміки маси тіла відрізняються від результатів інших досліджень [131], що, ймовірно, зумовлено низкою причин, як-то: видом розплідника, лінією самиць, їх більшою початковою вагою, сезоном року тощо і підкреслює, таким чином, важливість формування ідентичних контрольних та дослідних груп при проведенні експерименту [138].
Динаміка щотижневого приросту маси тіла інтактних самиць свідчить про його рівномірне збільшення протягом усього терміну вагітності з поступовим зростанням маси залежно від терміну — з 13,5±6,03 г на 7 день вагітності до 31,00±5,63 г — на 20 день вагітності, тобто у 2,3 рази (р<0,05). При цьому величина приросту маси тіла за останній тиждень вагітності (31,00±5,63 г) практично відповідає сумарному приросту 2-х попередніх тижнів (28,63±4,99 г), що цілком закономірно відображає період інтенсивного органогенезу у плодів, який починається з 9-го дня [78], збільшення розмірів плацент, а також підготовку організму вагітної самки до майбутніх пологів.
Таблиця 7.2.
Динаміка маси тіла (г) експериментальних тварин у різні терміни вагітності (M±m).
№ групи. | При; ріст. | Термін експерименту, дні (тижні). | |||||
|
|
|
|
| |||
1 (Pbа). | абс., г. | 12,25±6,32. | 11,25±6,94. | 19,75±7,13. | 23,50±6,32. | 42,25±6,42*. | |
%. | 106,56±3,47. | 105,66±3,59. | 109,40±3,5. | 112,59±3,56. | 122,64±3,7*. | ||
2 (Znх). | абс., г. | 9,63±5,82. | 14,38±7,76. | 22,88±9,77. | 24,00±7,62. | 46,88±8,31. | |
%. | 105,19±3,22. | 107,37±4,07. | 110,93±4,9. | 112,95±4,26. | 125,29±4,77. | ||
3 (Znц). | абс., г. | 14,63±4,60. | 15,75±4,89. | 24,13±6,58. | 30,38±4,98. | 54,50±6,37. | |
%. | 107,79±2,55. | 107,79±2,5. | 111,07±3,13. | 116,19±2,85. | 129,05±3,69. | ||
4 (Pbа+Znх). | абс., г. | 14,13±5,25. | 20,63±4,97. | 22,25±5,89. | 34,75±5,17. | 57,0±6,13. | |
%. | 107,71±2,99. | 110,45±2,66. | 110,21±2,8. | 118,96±3,13. | 131,11±3,79. | ||
5 (Pbа+Znц). | абс., г. | 9,50±4,52. | 15,50±5,57. | 26,38±8,85. | 25,00±5,26. | 51,38±8,23. | |
%. | 105,00±2,43. | 107,77±2,88. | 112,27±4,25. | 113,17±2,89. | 127,06±4,5. | ||
6 (К). | абс., г. | 13,50±6,03. | 15,13±5,63. | 31,00±5,63. | 28,63±4,99. | 59,63±6,03. | |
%. | 107,29±3,35. | 107,61±2,98. | 114,49±2,76. | 115,45±2,95. | 132,19±3,75. | ||
7 (Pbц). | абс., г. | 12,13±3,98. | 15,88±4,53. | 30,5±8,72. | 28,00±4,99. | 58,5±8,45. | |
%. | 106,55±2,24. | 108,05±2,35. | 114,31±4,21. | 115,12±2,87. | 131,6±4,79. | ||
Примітка. * - р<0,05 по відношенню до контролю.
Порівняння результатів динаміки маси тіла контрольної та дослідних груп виявило певні відмінності. Так, у тварин усіх дослідних груп спостерігалося різного ступеня зниження як абсолютних, так і відносних показників приросту маси тіла порівняно з інтактними самицями, проте, за винятком 1-ї дослідної групи, усі вони виявились невірогідними. Приріст маси тіла в самиць групи № 1 виявився у 1,4 рази (р<0,05) нижчим порівняно з контрольною групою, що може бути зумовлено як загально токсичним впливом свинцю на організм вагітної самки, так і його ембріотоксичністю. Аналогічна ситуація зменшення приросту маси тіла в цій дослідній групі порівняно з контрольною спостерігається в усі періоди вагітності, особливо на 2-му та 3-му тижнях. Така ситуація, ймовірно, пов’язана зі збільшенням загальної ембріональної смертності в дослідній групі № 1, як передімплантаційної (з 4−5 дня вагітності) [78], що проявляється зменшенням приросту маси тіла самиць на 2-му тижні, так і постімплантаційної, що відображається у динаміці приросту маси на 3-му тижні вагітності [27].
Аналіз динаміки краніокаудальних розмірів самиць щурів (табл. 7.3) свідчить про збільшення даного показника за період вагітності в усіх групах тварин у середньому на 1,35−1,73 см, що складає 7,74−9,86% від початкового розміру. При цьому відмінності ву динаміці розмірів тіла вагітних самиць дослідних та контрольної груп за весь період експерименту відсутні.
Проте якщо проаналізувати збільшення розмірів тіла самиць за окремі періоди вагітності, то помітно, що у контрольній групі, на відміну від усіх експериментальних груп, абсолютний приріст маси тіла за весь період вагітності в 1,7 разів (1,46 г проти 0,86 г), відносний — у 1,1 рази (108,3 проти 104,8) вищий порівняно з попередніми 2-ма тижнями вагітності (р<0,05), що закономірно відображає фізіологічні особливості збільшення організму тварин під час вагітності, більш виражені в останньому триместрі. Лише самиці дослідних груп № 3 та № 7, які отримували цитрати металів ізольовано, мають аналогічну тенденцію до зростання, що може свідчити про менш негативний їх вплив порівняно з неорганічними формами металів на перебіг вагітності.
Таблиця 7.3.
Динаміка довжини тіла (см) експериментальних тварин.
у різні терміни вагітності (M±m).
№ групи. | При; ріст, см. | Термін експерименту, дні. | |||||
|
|
|
|
| |||
1 (Pbа). | абс. | 0,41±0,39. | 0,45±0,32. | 0,53±0,31. | 0,86±0,34. | 1,39±0,39. | |
%. | 102,32±2,25. | 102,48±1,77. | 102,82±1,66. | 104,86±1,98. | 107,82±2,27. | ||
2 (Znх). | абс. | 0,27±0,24. | 0,57±0,31. | 0,59±0,40. | 0,85±0,27. | 1,44±0,34. | |
%. | 101,57±1,37. | 103,24±1,77. | 103,21±2,20. | 104,86±1,58. | 108,23±2,00. | ||
3 (Znц). | абс. | 0,43±0,33. | 0,39±0,38. | 0,66±0,38. | 0,81±0,33. | 1,48±0,33. | |
%. | 102,4±1,89. | 102,14±2,09. | 103,58±2,06. | 104,59±1,90. | 108,33±0,93. | ||
4 (Pbа+Znх). | абс. | 0,57±0,30. | 0,28±0,32. | 0,50±0,25. | 0,85±0,22. | 1,35±0,23. | |
%. | 103,30±1,71. | 101,53±1,78. | 102,73±1,40. | 104,87±1,27. | 107,74±1,34. | ||
5 (Pbа+Znц). | абс. | 0,38±0,33. | 0,53±0,41. | 0,82±0,41. | 0,90±0,38. | 1,73±0,32. | |
%. | 102,14±1,91. | 102,94±2,35. | 104,48±2,27. | 105,14±2,18. | 109,86±1,91. | ||
6 (К). | абс. | 0,49±0,17. | 0,37±0,18. | 0,60±0,17. | 0,86±0,20. | 1,46±0,16*. | |
%. | 102,77±0,98. | 102,07±1,0. | 103,25±0,96. | 104,8±1,14. | 108,3±0,97*. | ||
7 (Pbц). | абс. | 0,45±0,26. | 0,38±0,25. | 0,65±0,27. | 0,82±0,27. | 1,48±0,28. | |
%. | 102,54±1,51. | 102,07±1,41. | 103,51±1,49. | 104,66±1,58. | 108,34±1,66. | ||
Примітка. * - р<0,05 по відношенню до рівня приросту за перші 2 тижні.
Результати макроскопічних досліджень свідчать про відсутність відмінностей у зовнішньому вигляді внутрішніх органів інтактних та експонованих тварин. Відпрепаровані від клітковини яєчники жовтаво-рожевого кольору, округло-овальної форми з горбистою поверхнею, без помітних патоморфологічних змін. В яєчниках при огляді в бінокулярну лупу під невеликим збільшенням добре помітні жовті тіла (рис. 7.1) у вигляді виноградоподібних утворень рожевого чи жовтавого кольору, що дозволило провести кількісний їх аналіз для подальшого розрахунку гонадотоксичної дії досліджуваних металів.
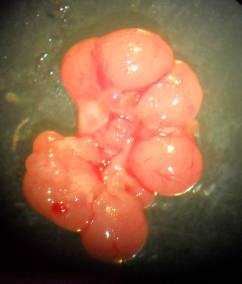
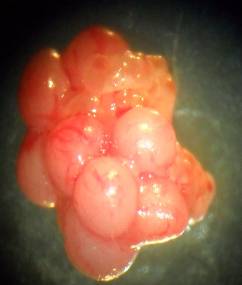
Рис. 7.1. Мікрофотографія правого (А) та лівого (В) незабарвленого нефіксованого яєчника вагітної самиці щура контрольної групи. Вирізняються жовті тіла (великого діаметру) та незрілі фолікули
(малого діаметру). Зб. 7×8.
Матка двороздільна, в кожному розі матки помітно від 1 до 8 ембріонів, кількість яких суттєво відрізнялась у контрольній та дослідних групах, особливо в групі, що отримувала ацетат свинцю (рис. 7.2). Місця імплантації плодів добре помітні, представляють собою білуваті ущільнення, злегка підняті над слизовою стінок рогів матки, відносно рівномірно віддалені одне від одного. У деяких випадках як у контрольній, так і в експериментальних групах у центрі місць імплантації помітно розширені кров’яні судини. Резорбовані плоди, що загинули на ранніх стадіях розвитку, мали вигляд невеликих темних ущільнень, на більш пізніх етапах — гомогенних тіл діаметром понад 5 мм.
Якщо макроскопічний огляд внутрішніх органів не виявив суттєвих відмінностей у розташуванні, формі та зовнішньому їх вигляді у тварин інтактних та експонованих груп, то визначення їх маси показало наявність певних особливостей (табл. 7.4). Так, маса серця та правої нирки у дослідній групі, що отримувала цитрат цинку ізольовано (група № 3) за середніми значеннями виявилась на 15,4% та 10,5% вищою, а маса серця у групі, яка отримувала цитрат свинцю, навпаки, — на 9% нижчою порівняно з контрольної групою (р<0,05).
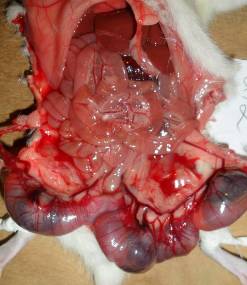
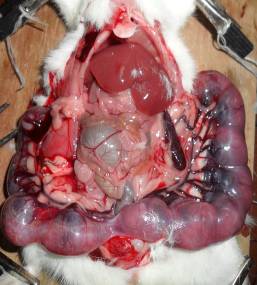
Рис. 7.2. Фотографія двороздільної матки самиці щура контрольної (А) та експериментальної (В) груп під час оперування. Виділяються маткові судини, що утворюють аркади в брижах маткових труб. Ембріони в маткових трубах вказано стрілкою
Звертає на себе увагу той факт, що середня маса печінки у дослідних тварин, які отримували ацетат свинцю ізольовано та у комбінації з хлоридом цинку виявилась на 12,5−23,2% вищою (р<0,05) порівняно з інтактними тваринами, що може бути зумовлено їх токсичним впливом на клітини печінки. У той же час, маса селезінки в групах № 2 і № 5 була на 17,1−20,3% нижчою (р<0,05) порівняно з групою контролю, що певним чином характеризує порушення постачання крові до даного органу. Маса правої нирки в середньому в усіх дослідних групах виявилась на 2−5 г вищою порівняно із масою лівої нирки, що співпадає з результатами досліджень інших учених [122] та свідчить про домінуючу роль правої нирки у виконанні видільної функції під час вагітності.
Абсолютна маса лівого яєчника самиць щурів дослідної групи № 1 виявилась на 30,8% (табл. 7.5) нижчою порівняно із контрольною групою (р<0,05). Сума мас правого та лівого яєчників у інтактних тварин становила 80,13±4,84 мг, що на 17,5% і 14,7% вище (р<0,05) порівняно з дослідними групами, які отримували ацетат свинцю ізольовано та у комбінації з хлоридом.
Таблиця 7.4.
Маса внутрішніх органів вагітних самиць щурів контрольної та дослідної груп (М±m).
№. групи. | Маса внутрішніх органів, г. | ||||||
серце. | печінка. | селезінка. | нирка права. | нирка ліва. | мозок. | ||
1 (Pbа). | 0,77±0,02. | 9,60±0,38*. | 1,5±0,13. | 0,78±0,01. | 0,76±0,02. | 1,66±0,04. | |
2 (Znх). | 0,81±0,03. | 9,29±0,48. | 1,02±0,03*. | 0,80±0,03. | 0,77±0,02. | 1,65±0,01. | |
3 (Znц). | 0,90±0,04*. | 8,75±0,40. | 1,14±0,04. | 0,84±0,02*. | 0,79±0,02. | 1,64±0,03. | |
4 (Pbа+Znх). | 0,86±0,05. | 10,51±0,89*. | 1,16±0,13. | 0,82±0,05. | 0,78±0,04. | 1,65±0,03. | |
5 (Pbа+Znц). | 0,79±0,04. | 8,05±0,37. | 0,98±0,04*. | 0,80±0,01. | 0,79±0,02. | 1,61±0,03. | |
6 (К). | 0,78±0,03. | 8,53±0,27. | 1,23±0,09. | 0,76±0,03. | 0,74±0,04. | 1,68±0,03. | |
7 (Pbц). | 0,71±0,02*. | 8,58±0,46. | 1,11±0,13. | 0,75±0,04. | 0,71±0,04. | 1,68±0,03. | |
Примітка. * - р<0,05 порівняно із контролем.
цинку. Отримані дані свідчать про негативний вплив сполук свинцю на масу яєчників, ступінь якого дещо нівелюється введенням цинку. У той же час сумарна маса яєчників у дослідній групі, що отримувала наноцитрат цинку ізольовано становить 81,25±3,25 мг, що вище контрольної групи та може свідчити про певну стимуляцію розвитку яєчників.
Комбіноване введення цитрату цинку з ацетатом свинцю (група № 5) призводить до вірогідного збільшення маси яєчників на 16% (р<0,01) порівняно з ізольованим введенням свинцю (група № 1) — 78,75±2,72 мг, що вказує на біопротекторну дію наноцинку. Порівняння характеру впливу неорганічних та органічних форм металів на масу яєчників виявило вірогідну відмінність між масою яєчників у дослідних групах № 2 та № 3 з вищими на 14,1% (р<0,05) показниками при введенні цитрату цинку [16].
Практично для усіх дослідних груп характерно пригнічення розвитку гонад за індексом впливу (І) досліджуваних сполук на масу яєчників, більш виражене для експериментальних груп, що отримували ацетат свинцю і хлорид цинку ізольовано та у їх комбінації (І = 0,83−0,87 ум.од.). Введення цитрату свинцю в організм вагітної самиці щура призводить до менш вираженої негативної дії на масометричні показники яєчників порівняно з його неорганічною формою (І = 0,93 та 0,83 ум.од. відповідно).
Таблиця 7.5.
Маса яєчників вагітних самиць щурів контрольної та дослідної груп (M±m).
№ групи. | Абсолютна маса, мг. | Сума масс правого та лівого яєчників, мг. | Індекс впливу (І),. ум. од. | ||
правий яєчник. | лівий яєчник. | ||||
1 (Pbа). | 39,57±7,59. | 26,57±1,72*. | 66,14±4,66*. | 0,83. | |
2 (Znх). | 36,63±6,76. | 33,17±2,72. | 69,80±4,74. | 0,87. | |
3 (Znц). | 47,25±3,58. | 34,00±2,92. | 81,25±3,25. | 1,01. | |
4 (Pbа+Znх). | 37,25±2,79. | 31,13±2,52. | 68,38±2,65*. | 0,85. | |
5 (Pbа+Znц). | 44,50±3,05. | 34,25±2,39. | 78,75±2,72**. | 0,98. | |
6 (К). | 41,75±6,23. | 38,38±3,45. | 80,13±4,84. | ; | |
7 (Pbц). | 38,88±7,03. | 35,38±4,11. | 74,25±5,57. | 0,93. | |
Примітки: * - р<0,05 по відношенню до контролю; ** - р<0,01 по відношенню до групи № 1.
Аналіз масометричних показників розвитку плаценти контрольної та дослідних груп (табл. 7.6, додаток М) свідчить, що її середня маса в усіх групах піддослідних тварин коливалася в межах 0,43−0,59 г, що становить 1,40±0,02−1,58±0,03% маси вагітних самиць та відповідає даним літератури [93].
Відмінності в масі плацент, як абсолютної, так і відносної, виявились невірогідними, за винятком дослідної групи, що отримувала наноцитрат свинцю (рис. 7.3), маса плаценти у якій на 27,1% нижча порівняно з контрольною групою (р<0,001). Аналогічна ситуація спостерігається і стосовно діаметру плаценти, який у даній дослідній групі на 7,3% (р<0,05) нижчий порівняно з інтактними тваринами — 1,40±0,02 см проти 1,51±0,04 см. Отримані дані свідчать про виражену негативну дію органічної форми свинцю на плацентогенез, що знайшло підтвердження при гістологічних та гістоморфометричних дослідженнях.

Рис. 7.3. Фотографія плацент самиць щурів дослідних груп, що отримували ацетат свинцю (А) та цитрат свинцю (В) після фіксації у формаліні
Незважаючи на те, що макроскопічні та масометричні показники розвитку плацент усіх груп виявились практично однаковими, результати гістологічного дослідження свідчать про розвиток дистрофічно-дегенеративних та дисциркуляторних змін структури усіх зон плаценти різного ступеня вираженості при збереженні загального плану будови органу.
Гістологічне дослідження плаценти самиці щура контрольної групи на 20-у добу вагітності показало, що сформована гемохоріальна плацента представлена трьома зонами (рис. 7.4): лабіринтна, спонгіотрофобластна та децидуальна зони. Найбільша за товщиною (1945,7±184,2 мкм) лабіринтна зона плаценти була утворена добре розвинутими анастомозуючими плодовими капілярами, трьома шарами клітин трофобластичного епітелію та лакунами, заповненими материнською кров’ю. Трофобластичний епітелій складався з більш розвинутого шару, оберненого в бік материнської лакуни, симпластотрофобласту, в якому виявлялися скупчення овальних і округлих ядер, а також направленого до фетальної судини менш виразного цитотрофобласту, що був представлений декількома рядами великих, витягнутої форми клітин з видовженими ядрами.
Зона спонгіотрофобласту складалась з компактно розташованих добре розвинутих клітин власне спонгіотрофобласта, глікогенних клітин, шару гігантських клітин трофобласта, материнських лакун та синусоїдів, заповнених кров’ю. Полігональні спонгіотрофобластичні клітини, в цитоплазмі яких виявлялися два або одне ядро з рівномірно розподіленим гетерохроматином, формували тяжі. Серед останніх поодиноко чи у формі кластерів розташовувались глікогенні клітини. Шар гігантських клітин трофобласта відділяв материнську частину плаценти від плодової. Серед них зустрічалися некротичні та дегенеруючі форми гігантських клітин спонгіотрофобласта, що були оточені переважно скупченнями нейтрофільних гранулоцитів.
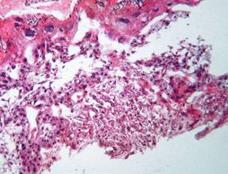
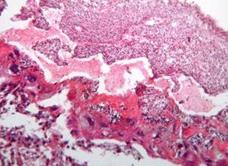
Рис. 7.4. Плацента щура контрольної групи. Лабіринтна та спонгіотрофобластична зони (А). Спонгіотрофобластична та децидуальна зони (В). Забарвлення гематоксиліном-еозином. Ч200
Децидуальна зона розташовувалася на периферії плацентарної тканини і була представлена скупченнями великих сплощених децидуальних клітин зі світлою цитоплазмою та овальними ядрами, що накопичували ліпіди і глікоген. Поміж них виявлялися лімфоцити та макрофаги. Хоріальна пластинка, яка розташовувалась з боку плодової частини плаценти, була не завжди добре помітна та утворювалась волокнистою тканиною з великими повнокровними судинами.
Про структурну зрілість плаценти свідчила наявність острівців глікогенних клітин у лабіринтному відділі, трансформація базофільних клітин у глікогенні, переважання питомого об'єму лабіринтного відділу над базальним, а також зближенням плодового та материнського кровотоків за рахунок витончення плацентарного бар'єру.
Введення ацетату свинцю зумовило розвиток дегенеративно-дистрофічних змін у плаценті (рис. 7.5). Незважаючи на збереження загальної товщини плаценти — 2867,5±185,4 мкм та структури органу, в усіх зонах плаценти визначалися патологічні зміни, характер яких залежав від досліджуваної ділянки. Товщина лабіринтної зони, що являє собою власне гемато-плацентарний бар'єр [171], перевищувала норму на 28,3% (p<0,05) і складала 2496,4±203,7 мкм. У цій зоні плаценти спостерігались ознаки деструктивно-дегенеративних процесів і набряку. Однак кількість плодових капілярів та ступінь їх розвитку знижувалися порівняно з контролем. Судини містили малу кількість формених елементів крові й місцями були порожніми, що, ймовірно, було пов’язано з погіршенням стану материнсько-плодового обміну. Також нерідко у капілярах спостерігалося явище сладж-синдрому еритроцитів. Для розширених материнських лакун було характерне застійне повнокров’я з формуванням стаза еритроцитів та тромбозу. Аналогічні зміни гістоструктури лабіринтної зони плаценти у щурів під впливом хімічних факторів довкілля виявлені й в інших дослідженнях [194, 209].
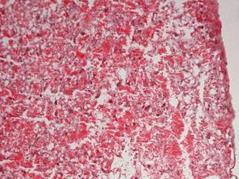
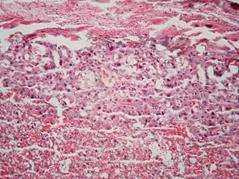
Рис. 7.5. Плацента самиці щура групи № 1. Лабіринтна зона (А). Спонгіотрофобластична та децидуальна зони (В). Забарвлення гематоксиліном-еозином. Ч200
У трофобласті лабіринтної зони виявлялося значне накопичення депозитів фібрину навколо ворсин порівняно з контролем, а їх мікроворсини були коротшими та менш розвиненими. Відзначалося порушення будови цитота синцитіотрофобластних балок лабіринту. У деяких ділянках цитотрофобласта, поряд із дистрофічними, виявлялися також некротичні зміни. У синцитії на місці накопичення гранул глікогену утворювались вакуолеподібні порожнини, відзначалося просвітлення його цитоплазми, вогнищеве скупчення ядер.
Товщина спонгіотрофобласту щурів складала 221,8±52,9 мкм, що було на 50,9% (p<0,05) нижче норми. У спонгіозній зоні спостерігалися гідропічна дистрофія спонгіотрофобластів, гіперхроматоз і руйнування їх ядер, дегенеративні зміни неглікогенних клітин, вогнища некрозу, крововиливів та місцями набряк строми. У цих місцях відзначалося також посилення руйнування в острівках глікогенних клітин, на місцях яких утворювались порожнини. Кількість гранул глікогену в них була нижчою за контроль. Шар гігантських клітин трофобласту був значно витончений та складався переважно з одного ряду дрібних клітин, у цитоплазмі яких спостерігались численні включення, що свідчило про збільшення фагоцитарної активності клітин. Площа крайових материнських синусоїдів та периферійних материнських лакун зменшувалась, в їх просвіті спостерігались явища тромбозу й агрегації еритроцитів.
Світлооптичне дослідження плаценти щурів експериментальних груп, які отримували сполуки цинку ізольовано показало, що загальний план будови органу і товщина гемохоріальної плаценти не відрізнялись від показників контрольної групи. Подібна ситуація характерна і для товщини лабіринтної зони плаценти при введенні хлориду цинку. У той же час введення органічної форми металу призвело до зниження даного показника на 24,4% (p<0,05).
У обох випадках лабіринтна зона була представлена добре сформованими плодовими капілярами в оточенні тришарового трофобласту та помірно розвинутими материнськими лакунами (рис. 7.6, 7.7). Витягнутої форми з овальним ядром клітини цитотрофобласту рівномірно вкривали фетальні судини, порушень в будові симпластотрофобластних білок виявлено не було.
Зона спонгіотрофобласту при введенні хлориду цинку була достатньо розвинута, за клітинним складом не відрізнялась від контрольної групи (рис. 7.6) і свідчила про її структурну зрілість.
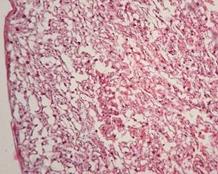
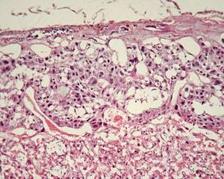
Рис. 7.6. Плацента щура групи № 2. Лабіринтна зона (А). Спонгіотрофобластична та децидуальна зони (В). Забарвлення гематоксиліном-еозином. Ч200
Ввведення цитрату цинку призвело до збільшення на 85,8% (р<0,05) товщини спонгіотрофобласту порівняно з контролем. Спонгіотрофобластичні клітини утворювали добре сформовані тяжі (рис. 7.7). Серед них відмічались великі скупчення глікогенних клітин, питома кількість яких значно перевищувала норму. У клітинах виявлялися ознаки фагоцитарної активності.
В усіх зонах плаценти при введенні комбінації ацетат свинцю-хлорид цинку виявлено різного ступеня виразності патологічні зміни. Загальний план будови органу зберігався, проте морфологічні особливості лабіринтної зони плаценти характеризувались деструктивно-дегенеративними процесами і гемодинамічними порушеннями, але ступінь їх вираженості був меншим порівняно з групою № 1 (рис. 7.8). Товщина лабіринтної зони (2075,1±207,6 мкм) дещо перевищувала аналогічний показник у контрольній групі за рахунок підвищеного кровонаповнення материнських лакун, проте без вірогідних відмінностей. Плодові капіляри були дещо спустошені, місцями в них виявлялись.
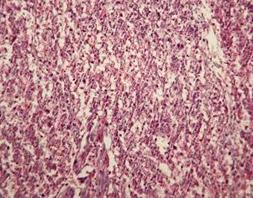
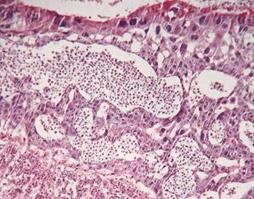
Рис. 7.7. Плацента щура групи № 3. Лабіринтна зона (А). Зона спонгіотрофобласту (В). Забарвлення гематоксиліном-еозином. Ч200
явища сладж-синдрому еритроцитів. У клітинах трофобласту виявлялися дистрофічні зміни, осередкове скупченням ядер синцитіотрофобласту в балках, накопичення депозитів фібрину. Товщина балок трофобласту значно варіювала. Окрім дистрофічних змін у них виявлялися некротичні вогнища. Питома кількість глікогенних клітин відповідала нормальним значенням. Материнські лакуни були помірно розширені, в них спостерігалось явище стазу еритроцитів, на периферії лабіринту — застійне повнокров’я, тромбоутворення.
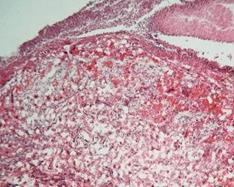
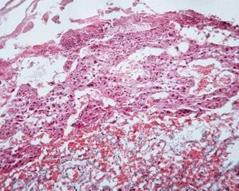
Рис. 7.8. Плацента щура групи № 4. Лабіринтна зона (А). Спонгіотрофобластична та децидуальна зони (В). Забарвлення гематоксиліном-еозином. Ч200
Товщина спонгіотрофобласту відставала від норми на 38,2% (p<0,05) та становила 279,7±56,8 мкм. Кількість спонгіотрофобластичних клітин була знижена, вони розташовувалися пухко, формуючи порожнини та розриви, часто заповнені фібриноїдом. Також відзначалось посилення руйнування глікогенних клітин, але воно було менш виразне, ніж у тварин групи № 1. Периферійні материнські лакуни і синусоїди були розширені, повнокровні та характеризувалися помірними ознаками застою і агрегації еритроцитів, в їх просвіті спостерігались явища тромбозу. У децидуальній зоні виявлялись помірні явища некрозу з лімфоцитарною інфільтрацією.
Аналіз лабіринтної зони експериментальної групи, що отримувала комбінацію ацетат свинцю-цитрат цинку (рис. 7.9) показав, що найбільш виразними були зміни структури трофобласту. Будова синцитію та цитотрофобластних балок характеризувалась неоднорідностю їх товщини, особливо поблизу фетальних судин. Кількість гігантських клітин трофобласту з великим щільним ядром перевищувала норму. У деяких ділянках цитотрофобласту спостерігались також некротичні вогнища, але їх кількість була незначна. Також зустрічались острівці глікогенних клітин. Ступінь розвитку фетальних капілярів не відрізнявся від групи контролю.
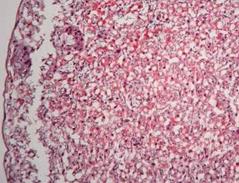
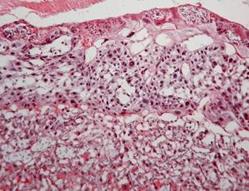
Рис. 7.9. Плацента щура групи № 5. Лабіринтна зона (А). Спонгіотрофобластична та децидуальна зони (В). Забарвлення гематоксиліном-еозином. Ч200
Показник товщини спонгіотрофобласту не відрізнявся від норми та складав 386,6±49,7 мкм. Ця зона була представлена полігональними спонгіотрофобластичними клітинами, що формували правильні тяжі. Серед них виявлялись скупчення глікогенних клітин. Шар гігантських клітин трофобласту характеризувався нормальним розвитком. Клітини розташовувались в декілька рядів та мали притаманну їм витягнуту, бобоподібну форму, овальне або округле ядро. Площа периферійних материнських лакун та крайових синусоїдів не відрізнялась від показників контрольної групи, у них спостерігалась помірна агрегація еритроцитів та зустрічались скупчення глікогенних клітин.
Гістоморфометричне дослідження плаценти щурів, які отримували цитрат свинцю протягом вагітності, показало, що в усіх зонах плаценти виявлялись дистрофічно-ішемічні зміни. Як загальна товщина плаценти, так і товщина лабіринтної зони були на 19,4−22,1% (р<0,01) нижчими за показники контрольної групи. Лабіринтна зона була представлена слабко розвиненими фетальними капілярами, кількість яких була значно знижена (рис. 7.10). Судини мали малу кількість формених елементів, були спустошеними та місцями зовсім спадалися. У трофобласті лабіринтної зони виявлялося значне накопичення депозитів фібрину навколо ворсин, мікроворсини були слабко розвиненими. Товщина цитота синцитіотрофобластних балок лабіринту була стоншена. У деяких ділянках цитотрофобласта поряд із дистрофічними виявлялися також некротичні зміни. Кількість гігантських клітин лабіринту з великим щільним ядром була знижена. Вищеперераховані зміни є наслідком вираженої ішемії та кисневого голодування й свідчать про зниження трансплацентарного транспорту в даній зоні, що узгоджується з результатами інших досліджень [70, 210].
Товщина спонгіотрофобласту виявилась на 49,5% (p<0,05) меншою за норму. У спонгіозній зоні спостерігалися гідропічна дистрофія спонгіотрофобластів, гіперхроматоз та руйнування їх ядер. Кількість спонгіотрофобластичних клітин була знижена, вони розташовувалися пухко, формуючи порожнини та розриви. Також відзначалось посилення руйнування глікогенних клітин, але воно було помірним, на їх місцях утворювались порожнини. Площа крайових материнських синусоїдів та периферійних материнських лакун зменшувалась. Серед скупчень лейкоцитів виявлялись некротично змінені та клітини з пікнотичними ядрами. Децидуальна зона характеризувалась збільшенням вогнищ некрозу та відкладанням депозитів фібрину. Питома кількість гранул ліпідів та питомий об'єм глікогену в децидуальних клітинах були різко знижені.
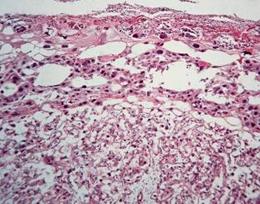
Рис. 7.10. Плацента щура групи № 7. Лабіринтна зона (А). Спонгіотрофобластична та децидуальна зони (В). Забарвлення гематоксиліном-еозином. Ч200
Вищеописані зміни структурної організації плаценти свідчать про дисциркуляторні порушення у ній, які в поєднанні з альтерацією трофо-бластичних клітин, складають, на нашу думку, патоморфологічний субстрат розвитку плацентарної дисфункції при дії сполук свинцю. В основі патогенезу даного процесу, ймовірно, лежить незавершена гестаційна перебудова материнських та фетальних капілярів, яка супроводжується зменшенням васкуляризації та розвитком дисциркуляторних розладів в плаценті. Суттєве зменшення питомого обсягу глікогенних клітин є додатковою ознакою структурної незрілості плацент. Крім того, зниження кількості глікогену в усіх зонах вказує на дефіцит енергетичного субстрату для забезпечення активного трансі параплацентарного транспорту, що узгоджується з результатами інших досліджень [209, 257, 318]. Циркуляторна гіпоксія приводить до пошкодження структури клітин трофобласту, а це, в свою чергу, веде до порушення транспортної, бар'єрної та трофічної функцій плаценти.
Завершуючи аналіз результатів вивчення впливу низьких доз ВМ на перебіг вагітності та плацентогенез, необхідною складовою повноцінної оцінки типу комбінованої дії бінарної суміші «свинець-цинк», на нашу думку, є розрахунок коефіцієнта їх комбінованої дії (Ккд).
Отримані результати (табл. 7.7) свідчать, що тип комбінованої дії свинцю та цинку за умови їх впливу на організм вагітної самиці щура у відносно низьких концентраціях, які не перевищують поріг загальнотоксичної дії, характеризується як антагоністичний практично для усіх досліджуваних фізіологічних, морфометричних та гістоморфометричних параметрів (Ккд=0,34−0,64). Виняток складає вплив комбінації вищезазначених металів на динаміку ректальної температури під час вагітності, який характеризується як синергічний (Ккд=1,52−1,63). На нашу думку, така ситуація зумовлена активізацією компенсаторних резервів організму вагітної самиці для забезпечення повноцінного формування та розвитку потомства в умовах подвійного навантаження ВМ, що потребує більше енерговитрат, а, отже, призводить до підвищення показників температури тіла. Проте, оскільки в усіх дослідних групах параметри ректальної температури не виходили за межі фізіологічних величин, даний синергічний вплив бінарної суміші «свинець-цинк» не є критичним та патогенетично значущим.
Ефект підгострої комбінованої дії свинцю і цинку ослаблений практично за усіма показниками в 1,6−3,0 рази, що співпадає з результатами подібних досліджень [162] та свідчить про зниження токсичного впливу свинцю в дозі 0,05 мг/кг на перебіг вагітності й плацентогенез за одночасного введення органічних та неорганічних сполук цинку в дозі 1,5 мг/кг. При цьому відмінностей у впливі різних форм цинку на масометричні параметри плаценти за їх комбінованої дії зі свинцем не виявлено. У той же час протекторна дія хлориду цинку по відношенню до токсичного впливу свинцю більш виражена при формуванні лабіринтної зони плаценти (Кзе=2,3), а цитрату цинкуспонгіотрофобластичної і децидуальної зон (Кзе=2,8−3,0), що співпадає з результатами гістоморфологічного дослідження.
Таблиця 7.7
Кількісна оцінка характеру комбінованої дії бінарної суміші.
«свинець-цинк» за показниками перебігу вагітності і плацентогенезу.
Параметри. | Коефіцієнт комбінованої дії (Ккд). | Кратність зміни ефекту (Кзе). | |||
Pbа+Znх. | Pbа+Znц. | Pbа+Znх. | Pbа+Znц. | ||
Динаміка маси тіла, г. | 0,64. | 0,53. | 1,6. | 1,9. | |
Динаміка довжини тіла, см. | 0,48. | 0,60. | 2,1. | 1,7. | |
Динаміка температури тіла, °С. | 1,52. | 1,63. | 0,7. | 0,6. | |
Маса плаценти, г. | 0,52. | 0,51. | 1,9. | 2,0. | |
Діаметр плаценти, см. | 0,53. | 0,51. | 1,9. | 2,0. | |
Товщина плаценти, мкм. | 0,45. | 0,49. | 2,2. | 2,1. | |
Товщина лабіринтної зони, мкм. | 0,44. | 0,54. | 2,3. | 1,9. | |
Товщина спонгіотрофобластичної зони, мкм. | 0,46. | 0,36. | 2,2. | 2,8. | |
Товщина децидуальної зони, мкм. | 0,64. | 0,34. | 1,6. | 3,0. | |
Експериментальні дослідження ембріотоксичності металів за ізольованої та комбінованої їх дії.
Результати вивчення ембріотоксичної дії низьких доз металів у неорганічній та органічній формах представлені в таблиці 7.8 (додаток М) і свідчать, що індекс плодовитості самиць у дослідних та контрольній групах практично збігався між собою і становив 0,8−0,9, що відповідає даним літератури [78]. При подальшому вивченні ембріотоксичної дії не враховували дві самиці з груп 1 та 3, оскільки в першому випадку відбулась резорбція усіх ембріонів на ранніх стадіях, а в другому випадку — смерть самиці внаслідок порушення внутрішньошлункового введеня препарату.
Показники загальної ембріональної смертності в контрольній групі тварин становили 11,11±4,43%, що знаходиться в межах спонтанного рівня, притаманного білим щурам та відповідає результатам досліджень інших авторів [78, 93, 108, 131, 236, 309], так само, як і показники дота постімплантаційної смертності - 0,10±0,05 од. і 0,01±0,01 од. відповідно. Введення ацетату свинцю протягом усього терміну вагітності призвело до збільшення ембріолетальності у 2,16 рази (р<0,01) порівняно з групою контролю, переважно в доімплантаційний період, що зумовило зниження на 17% кількості живих плодів на 1 самицю (р<0,05). Така ситуація підтверджує механізм регуляції чисельності плодів організмом самки на фоні впливу токсичної речовини, яка діє протягом усього періоду вагітності, в тому числі в доімплантаційний період (з 1 по 4−5 день вагітності). Енергетично для організму самиці щура більш раціонально абортувати плоди в початковий період вагітності, ніж у період інтенсивного органогенезу, що знайшло підтвердження і в інших дослідженнях з вивчення ембріотоксичності металів [108]. Згідно з даними [221], свинцю властива пряма токсична дія на гермінативний епітелій. Крім того, цей токсикант опосередковано зумовлює порушення взаємозв'язку між гіпоталамусом, гіпофізом і гонадами, призводить до підвищення збудливості мускулатури матки, що збільшує частоту виникнення викиднів та внутрішньоутробної загибелі плодів.
Введення цитрату свинцю призвело до менш вираженого ембріолетального ефекту — загальна ембріональна смертність становила 14,81±4,68%, що зумовило більшу чисельність плодів у посліді дослідної групи № 2 порівняно з введенням неорганічного свинцю — 8,63±0,80 проти 7,5±0,53 відповідно, при показниках у групі контролю — 9,0±0,4. Предімплантаційна смертність при введенні цитрату свинцю не відрізнялась від контрольної групи та була нижчою, ніж у групі, що отримувала ацетат свинцю. У той же час при введенні органічної форми металу спостерігалося збільшення у 1,5−3,0 рази показників постімплантаційної смертності порівняно з контрольною групою та групою № 1.
У дослідних групах, тварини яких отримували неорганічні та органічні сполуки цинку, встановлено відсутність вірогідних розбіжностей між рівнями загальної, дота постімплантаційної загибелі ембріонів, кількості живих плодів на 1 самицю порівняно з інтактною групою. До того ж в обох групах зазначені показники, з урахуванням похибки, знаходились у межах фізіологічного рівня, притаманного даному виду лабораторних тварин [92, 108, 119, 120].
У тварин експериментальних груп, які отримували комбінацію ацетат свинцю-хлорид цинку, спостерігалася тенденція до збільшення кількості живих плодів у посліді порівняно з групою № 1 — 8,75±0,4 (р=0,06), що зумовлено зниженою на 37,6% (р<0,001) загальною ембріональною смертністю. При цьому вищезазначені показники практично не відрізнялися від контрольної групи.
При комбінованому введені свинцю та цитрату цинку виявлено збільшення загальної кількості живих плодів у посліді на 23,3% (р<0,05) за рахунок зниження у 3,2 разів рівня загальної ембріональної смертності (р<0,001) порівняно з групою № 1.
Стосовно розподілу плодів за статтю у посліді, то у контрольній групі тварин спостерігалося 45,83±3,17% самців та 54,17±3,8 самиць, що знаходиться в межах видових значень [78, 93]. За умови впливу низьких доз свинцю як при ізольованому його введенні, так і у комбінації з неорганічними та органічними сполуками цинку таке співвідношення зберігалося, хоча характеризувалося деяким підвищенням питомої ваги осіб жіночої статі - 55,71−56,76% самиць проти 42,86−43,33% самців, що збігається з даними інших досліджень [119, 120, 205]. У той же час у групах, що отримували препарати цинку ізольовано, так само, як і при введенні цитрату свинцю, навпаки, спостерігалося деяке збільшення кількості самців у посліді - 49,28−53,85% проти 46,15−50,72% самиць, хоча, як і у попередньому випадку, ці відмінності невірогідні. Отримані дані свідчать про відсутність суттєвих змін у співвідношенні плодів чоловічої та жіночої статі у посліді при введенні низьких доз металів.
Показники маси, краніокаудальних розмірів та діаметру тіла плодів у експериментальних групах, тварини яких отримували свинець і цинк у неорганічній та органічній формах, вірогідно не відрізнялися від тварин контрольної групи за винятком тенденції до зниження на 3,3% краніокаудального розміру в дослідній групі № 1. Проте аналіз даних показників за статтю виявив зниження на 11,4% (р<0,01) діаметру тіла самиць у дослідній групі, яким вводили цитрат цинку ізольовано, а також зниження та 4,7% (р<0,05) краніокаудального розміру самців у групі, яка отримувала ацетат свинцю — 30,11±0,44 мм проти 31,6±0,46 мм у групі контролю, що збігається з результатами інших досліджень [221].
У той же час у групах, які отримували комбінацію ацетату свинцю і цинку (дослідні групи № 4, № 5), спостерігалося зниження маси та краніокаудальних розмірів усіх плодів на 3,7−11,4% (р<0,05). Такі відмінності у масометричних показниках зумовлені вірогідною різницею у плодів чоловічої статі - на 3,7- 11,9% порівняно з групою контролю (р<0,01). У даних групах спостерігалося зниження діаметру тіла плодів жіночої статі на 7,7% (група № 4), чоловічої статі - на 9,7% (група № 5) (р<0,01). Вищенаведені дані можуть свідчити про дещо більшу чутливість плодів чоловічої статі по відношенню до впливу низьких доз свинцю під час вагітності, що не нівелюється введенням його біоантагоністу — цинку і зумовлено, ймовірно, незрілістю захисних систем організму [37].
Що стосується відмінностей у масі плодів чоловічої та жіночої статі, то як у контрольній, так і в дослідних групах вони суттєво не відрізнялися. Виняток становить дослідна група № 3, де середня маса тіла плодів чоловічої статі виявилась на 11,3% (р<0,05) вищою порівняно з самками. У той же час в усіх групах, що отримували препарати цинку ізольовано, а також у комбінації ацетат свинцю-хлорид цинку, краніокаудальний розмір плодів чоловічої статі був на 2,5−6,2% вищим порівняно з плодами жіночої статі (р<0,05), що може свідчити про деяку стимуляцію розвитку самців за умови підвищеного надходження цинку до організму вагітної. Цей показник виявився найвищим у групі, тварини якої отримували цитрат цинку ізольовано — 31,67±0,23 мм у самців проти 29,71± 0,13 мм — у самок. Діаметр плодів чоловічої статі виявився на 4,3−16,2% (р<0,05-р<0,001) вищим порівняно з самицями в дослідних групах № 4 і № 5. У той же час діаметр тіла самиць у групах, які отримували ацетат свинцю ізольовано та у комбінації з цитратом цинку, був на 4,5−6,1% вищим (р<0,05) порівняно з діаметром тіла самців, що може свідчити про більш негативну внутрішньоутробну дію свинцю на розвиток плодів чоловічої статі.
Аналіз типу комбінованої дії бінарної суміші «свинець-цинк» за показниками ембріотоксичності (табл. 7.9) свідчить про антагонізм вищезазна-чених металів у відносно низьких дозах при впливі на ембріональний розвиток щурів (Ккд=0,19−0,75). При цьому, на відміну від результатів аналізу впливу різних форм цинку на прояви токсичності свинцю за показниками перебігу вагітності та розвитку плаценти, біопротекторна дія цитрату цинку по відношенню до ембріогенезу щурів виявилась більш виразною порівняно з неорганічною формою металу. Попри ідентичну здатність обох форм цинку знижувати негативний вплив свинцю на морфометричні параметри розвитку плодів (Ккд=0,46−0,50; Кзе=2,0−2,2), відзначається надзвичайно ефективна протекторна дія цитрату цинку по відношенню до токсичності свинцю за показниками ембріолетальності (Ккд=0,19−0,33; Кзе=3,0−5,3). У той же час введення хлориду цинку на фоні свинцевої інтоксикації ослаблює ембріотоксичну дію свинцю лише у 1,3−2,9 разів.
Таблиця 7.9.
Кількісна оцінка характеру комбінованої дії бінарної суміші.
«свинець-цинк» за показниками ембріотоксичності.
Параметри. | Коефіцієнт комбінованої дії (Ккд). | Кратність зміни ефекту (Кзе). | |||
Pbа+Znх. | Pbа+Znц. | Pbа+Znх. | Pbа+Znц. | ||
Кількість живих плодів на 1 самицю. | 0,39. | 0,59. | 1,8. | 1,7. | |
Загальна ембріональна смертність, %. | 0,39. | 0,19. | 2,6. | 5,3. | |
Предімплантаційна смертність, од. | 0,34. | 0,19. | 2,9. | 5,3. | |
Постімплантаційна смертність, од. | 0,75. | 0,33. | 1,3. | 3,0. | |
Маса тіла 1 плода, г. | 0,49. | 0,46. | 2,0. | 2,2. | |
Краніокаудальний розмір 1 плода, мм. | 0,50. | 0,49. | 2,0. | 2,1. | |
Діаметр 1плода, мм. | 0,48. | 0,48. | 2,1. | 2,1. | |
Для виявлення зрілості фетоплацентарного комплексу розраховано плодово-плацентарний коефіцієнт, який у контрольній та дослідних групах № 1-№ 5 знаходився в межах фізіологічних значень для даного періоду гестації - 0,23−0,27 ум.од. [171]. Виняток становить величина плодово-плацентарного коефіцієнту в дослідній групі, що отримувала цитрат свинцю — 0,17 ум.од., що зумовлено меншим розміром плацент. Згідно з даними [257], така величина коефіцієнту свідчить про зрілість фетоплацентарного комплексу і спостерігається наприкінці вагітності самиць щурів — на 21−22 день. У той же час у наших дослідженнях, при однаковому з іншими групами терміні вагітності, маса тіла плодів залишалася практично незмінною при значно нижчих морфометричних показниках плаценти. Даний факт може свідчити про порушення розвитку плаценти за умови впливу низьких доз свинцю, більш вираженому при введенні його у органічній формі. Головним патоморфологічним механізмом такого впливу може бути зниження об'єму та питомої ваги фетальних судин плаценти під дією свинцю, що супроводжується зниженням загальної маси й діаметру органу.
Масо-ростовий коефіцієнт в усіх дослідних та контрольній групах знаходився у межах 0,71−0,79 ум.од., що свідчить про збереження пропорційності розвитку плодів за умови впливу низьких доз досліджуваних металів.
При макроскопічному дослідженні ембріонів, резорбованих на різних стадіях, а також плодів усіх експериментальних груп виявили певні особливості, обумовлені терміном гестації та формою досліджуваного металу. Так, резорбовані ембріони, які загинули на ранніх стадіях розвитку, мали вигляд невеликих темних ущільнень, на більш пізніх етапах — гомогенних тіл діаметром понад 4 мм. При цьому видимих вад розвитку плодів контрольної та усіх дослідних груп не виявлено, за винятком незначної кількості підшкірних геморагій (рис. 7.11-А), кількість яких знаходилась у межах, притаманних даній групі тварин [78]. У той же час у дослідній групі № 1 на тілі деяких плодів виявлено білуваті утворення, подібні до кальцинатів, а також кісти (7.11-В).
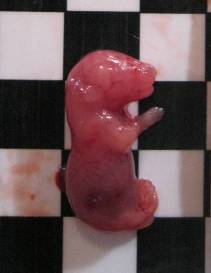
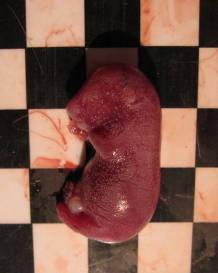
Рис. 7.11. Підшкірні геморагії (А), кальцинати та кісти (В) у плодів експериментальної групи № 1
Гігієнічна оцінка механізмів міграції мікроелементів та особливостей їх транслокації в системі «мати-плацента-плід».
Дослідження кількісного вмісту мікроелементів у крові інтактних вагітних тварин показало, що їх вміст, за середніми значеннями, коливався від 0,0048±0,0009 мг/кг для кадмію до 2,89±0,19 мг/кг — для цинку (табл. 7.10). Отримані нами дані щодо мікроелементного складу крові відповідають результатам інших досліджень [55] по відношенню до токсичних металів, у той час, як вміст міді виявився вищим у 1,9 разів, а цинку, навпаки, нижчий у 2,2 рази (р<0,05). Така ситуація може бути зумовлена низкою причин, серед яких — особливості хімічного складу харчових продуктів Дніпропетровської області, а також використання методів дослідження з різною чутливістю.
Вміст свинцю у крові вагітної самиці щура при введенні металу в органічній і неорганічній формах збільшується, відповідно, в 1,6 (р<0,01) і 1,8 разів (р<0,001) порівняно з контрольною групою тварин та співпадає з результатами інших досліджень [228, 363].
Слід зазначити, що при накопиченні свинцю в організмі відбуваються зміни елементного складу крові та порушення фізіологічного співвідношення есенціальних мікроелементів між собою. Так, при введенні сполук свинцю спостерігаються діаметрально протилежні зміни концентрацій цинку і міді в крові - на фоні активного зниження вмісту цинку в 2,9−3,1 рази суттєво збільшується концентрація міді - у 2,2−2,4 рази (рмідь>свинець>кадмій, то при введенні сполук свинцю даний ряд набуває нового вигляду: мідь>цинк>свинець>кадмій, тобто відбувається підвищення рівня міді за рахунок значного зниження концентрації цинку. При введенні свинцю різко змінюється співвідношення Cu: Zn, яке становить 1:0,5, у той час як за фізіологічного перебігу вагітності воно складає 1:3,3. Важливим є те, що таке співвідношення есенціальних мікроелементів спостерігається і у крові жінок [234], що підтверджує значущість екстраполяції експериментальних даних для розуміння механізмів міграції ВМ у організмі людини.
Таблиця 7.10.
Вміст важких металів у крові самиць щурів (M±m).
Груп… |