Збуджувальні та гальмівні нейромедіатори

Тіла дофамінергічних нейронів знаходяться в середньому мозку (чорна субстанція, вентральна покришка), нюховій цибулині, гіпоталамусі і паравентрикулярній області продовгуватого мозку. Дофамінергічні тракти з'єднують чорну субстанцію з неостріатумом, вентральну покришку з лімбічною системою і з лобною корою; n. arcuatis — ядро гіпоталамуса із туберо-інфундібулярною системою (серединне підвищення… Читати ще >
Збуджувальні та гальмівні нейромедіатори (реферат, курсова, диплом, контрольна)
Ацетилхолін: попередником служить холін, який потрапляє з їжею.

Ацетилхолін (АХ) широко представлений в різних відділах нервової системи. Синтез та розщеплення АХ відбувається в холінергічних структурах центральної та периферийної нервової системи. Основна його кількість зосереджена в периферийних нервово-м'язових синапсах, рецептори яких відносяться до категорії нікотинових (н-холінорецептори) — рецептори, які збуджуються нікотином. В ЦНС ацетилхолін зосереджений переважно в базальних гангліях, таламусі та сірій речовині. Відповідні рецептори в мозку відносяться до мускаринових (м-холінорецептори) — рецептори, які збуджуються мускарином — токсином грибів. АХ в підкоркових структурах приймає участь в регуляції складних рухливих функцій. Пошкодження холінергічної інервації в структурах мозку супроводжуються порушенням складних рухливих функцій. Так, наприклад, при паркінсонізмі поряд з порушенням дофамінергічної трансмісії відмічається гіперактивність деяких холінергічних систем мозку. Порушення холінергічної передачі в периферийній нервовій системі, особисто в нервово-м'язових синапсах, пов’язано з симптомами «втомленості» і «слабкості» м’язів. Припускають, що в основі тяжкого захворювання — міастенії гравіс (miastenia gravis) лежить аутоімунний процес, при якому антитіла блокують функцію холінорецепторів.
Холін вступає в холінергічні нейрони за допомогою спеціфічних систем транспорта. Синтез протікає в цитоплазмі клітин за участю холінацетилтрансферази (ХАТ).
Холін + Ацетил-КоА ХАТ ацетилхолін + КоА Потім АХ потрапляє в синаптичні везикули. Після екзоцитоза АХ в синаптичну щілину він підлягає інактивації за участю ацетилхолінестерази (АХЕ):
АХ АХЕ холін + ацетат АХ є переважно збуджуючим нейромедіатором, рідше — гальмівним (наприклад, в синапсах n. vagus). Холінергічні нейрони локалізуються в наступних відділах мозку: медіальному ядрі перетинки, діагональній зв’язці, базальному гігантоклітинному ядрі, ядрах мосту. Аксони цих нейронів мають проекцію на гіпокамп та проходять через кору великих півкуль. Холінергічні нейрони головного мозку приймають участь в таких функціях, як пам’ять, регуляція рухливості, рівні збудливості. Холінергічні синапси мозку вміщують переважно мускаринові рецептори. В спинному мозку АХ є нейромедіатором в синапсах, які утворюються б-мотонейронами на клітинах Реншоу.
В вегетативній нервовій системі АХ є нейромедіатором в усіх прегангліонарних нервових закінченнях симпатичної і парасимпатичної нервової системи (в тому числі і в мозковій речовині наднирників) через нікотинові холінорецептори. АХ здійснює через нікотинові холінорецептори нейромедіаторну функцію у нервово-м'язових синапсах, які утворені соматичними еферентними нервами в скелетних м’язах.
Моноаміни. До медіаторів цієї групи відносяться катехоламіни, а також серотонін і гістамін. В группу катехоламінів входять норадреналін (НА), адреналін (А), дофамін (ДА). Попередником катехоламінів є L-тирозин, який організм отримує з їжею, та здатний синтезуватися в печінці із амінокислоти фенілаланіна. L-Тирозин надходить в нервові закінчення шляхом активного транспорта, де протікає синтез катехоламінів по наступній схемі:
L-тирозин тирозингідроксилаза ДОФА декарбоксилаза дофамін.
дофамін-в-гідроксилаза норадреналін фенілетаноламін-N-метилтрансфераза адреналін Катехоламіни депонуються в крупних синаптичних везикулах (40−140нм). Розщеплення катехоламінів проходить за участю моноамінооксидаз і катехол-о-метилтрансферази. Інактивація синаптичного НА після його екзоцитоза здійснюється також шляхом зворотнього поглинання в нервові закінчення, тобто завдяки активного трансмембранного транспорта.
Норадреналін (НА):
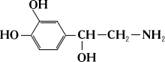
Тіла норадренергічних нейронів в ЦНС знаходяться в стволі мозку, головним чином в мосту мозку (голуба пляма, латеральна ретикулярна формація моста), в продовгуватому мозку і ядрі одиночного тракту. Багаточисленні нейрони голубої плями утворюють дифузні проекції, які досягають практично всіх відділів ЦНС — кори великих півкуль, лімбічної системи, таламуса, гіпоталамуса, спинного мозку. Низхідні норадренергічні шляхи спинного мозку приймають участь в регуляції м’язів-згибателів і судинного тонуса. В вегетативній нервовій системі НА є нейромедіатором постгангліонарних синаптичних нервів. У мозковій речовині наднирників вивільняється НА і А. В ЦНС НА є в ряді відділів переважно гальмівним нейромедітором, наприклад, в корі великих півкуль. В гіпоталамусі НА виконує збуджувальну нейромедіаторну функцію. Вважають, що моноамінові нейромедіатори мають відношення до прояву емоцій, регуляції настрою. Порушення обміну НА пов’язують із виникненням ряда психоемоційних розладів (шизофренія, маніакально-депресивних станів).
Адреналін (А):

В головному мозку кількість адренергічних шляхів більш обмежена в порівнянні з норадренергічними. Тіла нейронів, які містять фенілетаноламін-N-метилтрансферазу (фермент, який здійснює синтез А) знаходяться в нижніх відділах моста і в продовгуватому мозку. Низхідні шляхи досягають центральної сірої речовини і ядер гіпоталамуса (особисто, паравентрикулярного і дорсомедіального). Нейромедіаторна роль, А сумнівна. Нейромедіатором адренергічних нейронів є, очевидно, НА. Адреналін вивільняється дифузно (наприклад, разом з НА в мозковій речовині наднирників) і виконує роль модулятора.
Дофамін (ДА):
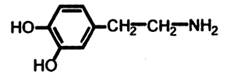
Тіла дофамінергічних нейронів знаходяться в середньому мозку (чорна субстанція, вентральна покришка), нюховій цибулині, гіпоталамусі і паравентрикулярній області продовгуватого мозку. Дофамінергічні тракти з'єднують чорну субстанцію з неостріатумом, вентральну покришку з лімбічною системою і з лобною корою; n. arcuatis — ядро гіпоталамуса із туберо-інфундібулярною системою (серединне підвищення). Дофамін має медіаторні властивості, бере участь у регуляції поведінки, діяльності серцево-судинної системи, кишечника, нирок. Дофамін служить нейромедіатором амакринових клітин сітчатки. У дофамінергічних нейронах аксони знаходяться в передньому мозку, де вони приймають участь у розвитку емоційних реакцій. Розрізняють декілька типів дофамінових рецепторів Д1, Д2, Д3, Д4. Важливу роль дофамінова медіаторна система відіграє в регуляції складних рухливих функцій. Порушення дофамінергічних шляхів призводить до скутності рухів, особливо стереотипних, до виникнення мимовільного дрижання і скутності м’язів, тобто характерних симптомів паркінсонізма. Гіперфункція дофамінергічної системи тісно пов’язана з механізмами розвитку шизофренії.
Серотонін (5- гідрокситриптамін, 5- HT):
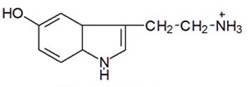
Попередником біогенного аміну 5- HT є незамінна амінокислота Lтриптофан. Синтез 5- HT протікає позасекреторними гранулами, які поглинають його за допомогою високоафінного переносія:
L-триптофан триптофангідроксилаза 5-гідрокситриптофан.
декарбоксилаза 5-гідрокси-триптамін.
Після екзоцитоза 5-HT інактивується шляхом активного зворотнього транспорту або підлягає окислювальному дезамінуванню з утворенням 5-гідроксиіндолоцтової кислоти, яка екскретується з сечею. Серотонінергічні нейрони входять до складу ядра шва в ростральній частині моста мозку. Ці нейрони дають проекцію на лімбічну систему, базальні ганглії, кору великих півкуль. В продовгуватому мозку знаходяться серотонінергічні нейрони, аксони яких утворюють шляхи в ствол мозку і спинний мозок. Крім того, серотонін виявлено в нейронах гіпоталамуса, чорній субстанції, спинному мозку. Відіграє важливу роль в регуляції емоційної поведінки, рухомої активності, терморегуляції, а також приймає участь в контролі нейроендокринної системи. Серотонін може виконувати функцію не тільки нейромедіатора але й нейромодулятора (нейрогормона). Лікарські засоби, які здатні частково знижувати виділення із синапсів моноамінові нейромедіатори (дофамін, норадреналін і серотонін), визивають депресію, тоді як всі препарати, що застосовуються для лікування клінічної депресії підвищують вміст цих нейромедіаторів в головному мозку та їх дію.
Найбільш розповсюдженними в тканині мозку нейромедіаторами є деякі амінокислоти. Вони розподіляються на дві групи: збуджувальні амінокислоти (глутамат, аспартат) і гальмівні (ГАМК, гліцин, таурин).
Глутамінова кислота:
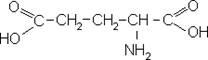
Попередником для синтеза глутамінової кислоти є глутамін, який поступає з глії в нейрон і є субстратом для синтеза глутамата, аспартата і ГАМК за схемою:
глутаміназа аспартатамінотрансаміназа глутаматдекарбоксилаза.
L-глутамінова кислота — основний збуджувальний нейромедіатор. Глутамат виявляється в усіх відділах ЦНС, очевидно тому, що він є не тільки нейромедіатором, але і попередником для синтезу інших амінокислот. Тіла глутаматергічних нейронів знаходяться в корі великих півкуль, нюхальний луковиці, гіпокампі, чорній субстанції, мозочку, сітківці. Глутаматергічні синапси існують в миндалині, стріатумі, на клітинах-зернах мозочка. В спинному мозку глутамат зосереджений в первинних аферентних волокнах дорзальних коришків. Глутаматергічні синапси розповсюдженні в корі головного мозку, гіпокампі, смугастому тілі і гіпоталамусі. Низхідні глутаматергічні шляхи виявлені практично в усіх структурах головного мозку, проекції яких йдуть від кори до підкоркових структур. Нейрорецептори глутамата розташовані кластерами на постсинаптичній мембрані. Порушення глутаматергічної медіації пов’язано з цілим рядом патологічних станів нервової системи: епілепсія, розлади вестибулярної системи, гіпоксиєю і ін.
Аспарагінова кислота:

Найбільш високий вміст аспартата знайдено в середньому мозку. В спинному мозку аспартат міститься у дорзальній і вентральній сірій речовині. Він приймає участь в збуджуванні інтернейронів, які регулюють різні спинномозкові рефлекси.
г-Аміномасляна кислота (ГАМК) представляє собою основний гальмівний нейромедіатор ЦНС:

У корі головного мозку приблизно в 5% нервових закінченнях виявляється ГАМК. Вона знаходиться в нейронах стріатума, які дають проекції на чорну субстанцію, гіпоталамусі, в нейронах мозочка (клітини Пуркін'є, корзинчаті та зіркові клітини). В желатинозній субстанції задніх рогів спинного мозку присутні ГАМКергічні аксо-аксональні синапси на первинних аферентних волокнах. Ці синапси опосередковують деполяризацію і гальмування секреції нейромедіатора — пресинаптичне гальмування. ГАМК приймає участь в регуляції моторної активності, підтримці судомного порога, формуванні емоційної поведінки. ГАМКергічна система приймає участь в здійсненні умовних рефлексів, організації процесів навчання і пам’яті. При цьому вона тісно взаємодіє з іншими медіаторними системами мозку: дофамінергічною, холінергічною і глутаматергічною. ГАМК-ергічна система може бути пов’язана з механізмами багатьох метаболічних розладів нервової системи. З порушеннями в цій системи пов’язані прояви епілепсії, хореї Гентингтона, паркінсонізма і деякі інші пошкодження екстрапірамідної системи.