Особливості будови мітохондрій скелетних м"язів при фізичному навантаженні в експерименті

Одночасно в ультраструктурі м’яза з’являються ділянки (~ 20%), що переповнені мітохондріями. Такі ділянки у тварин І підгрупи, де виражені компенсаторно-пристосувальні процеси, становлять > 50%, через що величина кількісної щільності суттєво (більш ніж удвічі) і достовірно перевищує середній показник у ІІ підгрупі. Відмінності мітохондрій за площею зрізу в І та ІІ пі-Оригінальні дослідження… Читати ще >
Особливості будови мітохондрій скелетних м"язів при фізичному навантаженні в експерименті (реферат, курсова, диплом, контрольна)
Особливості будови мітохондрій скелетних м’язів при фізичному навантаженні в експерименті.
Довготривала адаптація до значних фізичних навантажень є основою спортивних тренувань, фундаментом для досягнення високих спортивних результатів [1]. При багаторічних значних фізичних навантаженнях дуже важливим є своєчасне виявлення несприятливих порушень і вимагає глибокого вивчення даної проблеми. Зміни в процесі систематичної м’язової роботи мають пристосувальний характер і відбуваються на всіх рівнях структурної організації організму. Значні фізичні навантаження, особливо на тлі тривалого тренувального процесу, можуть викликати негативні зміни в організмі, призводити до напруження адаптаційних механізмів та їхнього зриву, що виявляється в порушеннях функціонального стану не тільки серцево-судинної та імунної систем [2, 3], які добре вивчені, але й інших життєво важливих систем організму. Фізичне навантаження під час тренувань і змагань є причиною прямого впливу на зміни структури м’язів та їхню функціональну активність.
Дослідження проведені у рамках НДР 2.24. «Підвищення ефективності тренувальної та змагальної діяльності кваліфікованих спортсменів дозволеними засобами відновлення та стимуляції працездатності» (номер держреєстрації 0111U001731) Зведеного плану науково-дослідних робіт у сфері фізичної культури і спорту Міністерства освіти і науки, молоді та спорту України на 2011;2015 рр.
Мета дослідження: вивчення морфофункціональних особливостей будови мітохондрій лит;
58 кового та камбалоподібного м’язів експериментальних тварин.
Матеріали і методи. Дослідження проводили на 20 білих статевозрілих щурах лінії Фішер з початковою масою 200−220 г. Тварин порівну було розподілено на контрольну (інтактні щури) та основну групу, в якій фізичне навантаження відтворювали шляхом застосовування протягом 5 тижнів щоденного плавання тварин по 30 хвилин з додатковим вантажем, що складав 10% від маси тіла. Утримання і використання лабораторних тварин відповідало методам, які рекомендуються національними нормами з біоетики [4]. Після декапітації під ефірним наркозом шматочки м 'язів фіксували 2,5%розчином глютарового альдегіду на фосфатному буфері з дофіксацією у 1% розчині чотириокису осмію за Мілонінгом. Зневоднювання проводили у спиртах зростаючої концентрації та ацетоні. Просочували та заливали у суміш епон-аралдит згідно із загальноприйнятою методикою [5].
Вивчення матеріалу литкових та камбалоподібних м 'язів щурів обох груп проводили на трансмісійному електронному мікроскопі ПЕМ-125К (Росія) з подальшим фотографуванням. Отриманий цифровий матеріал опрацьовували методами варіаційної статистики за допомогою прикладних пакетів комп 'ютерних програм «Statgmphics» і «Excel 97». Достовірність змін оцінювали з урахуванням t-критерію Ст 'юдента. [6].
Результати досліджень та їх обговорення.
Мітохондрії в литковому м’язі розташовуються підсарколемно, поміж саркомерами та розрізняються за формою та розмірами. Загалом їх кількість дорівнює (93,19± 16,03) х10-2 в мкм3, що значно більше, ніж у литковому м’язі інтактних тварин (табл. 1). При аналізі розподілу мітохондрій за цим показником видно, що таке збільшення відбувається внаслідок суттєвого зниження числа ділянок, які були притаманні інтактним щурам, та появі ділянок, майже повністю заповнених мітохондріями (рис. 1).
Як свідчать отримані дані (табл. 1), збільшується не тільки кількість, а й середня площа зрізу мітохондрій у порівнянні з інтактними тваринами. Це зумовлено появою великих за розмірами органел, яких не було у інтактних щурів, на тлі зменшення числа найдрібніших мітохондрій.
Слід зазначити, що такі органели у симпластах після фізичного навантаження розташовуються, як правило, поодиноко, поміж саркомерів (див. рис. 1).
Таблиця 1Морфометричні показники мітохондрій (МТ) у симпластах литкового м’яза щурів контрольної та основної груп
Групи тварин. | Показники, що вивчались (х ±S). | ||||
Об'ємна щільність МТ, %. | Кількісна щільність МТ, 10-2 х-3. мкм3 | Площа зрізу МТ, 10-2 мкм2 | Фактор форми. | ||
Контроль. (n=6). | 3,37±0,57. | 22,2±5,18. | 15,93±0,67. | 0,83±0,01. | |
Основна, в тому числі: | |||||
підгрупа І (n=3). | 27,37±7,17*. | 93,19±16,03*. | 43,68±1,88*. | 0,82±0,01. | |
підгрупа II (n=3). | 7,96±, 24* #. | 39,62±5,78#. | 30,18±2,01* #. | 0,75±0,01* #. | |
Примітка: * - статистично вірогідно порівняно з даними в контрольній групі; # - порівняно з даними в підгрупі І (р<0,05).
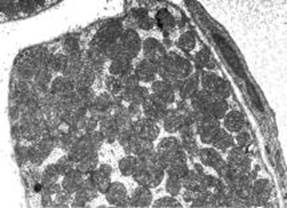
Рис. 1. М’язові волокна у литковому м’язі щурів підгрупи-! після довготривалого фізичного навантаження. Скупчення мітохондрій. Електронномікроскопічне фото. Зб.: 13 000.
мітохондрія м’язи фізичний навантаження Але здебільшого мітохондрії утворюють підсарколемні скупчення або ланцюжки поміж волокон, як це характерно для кардіоміоцитів. Серед мітохондрій у цій групі тварин переважають кулястої форми органели, що підтверджується показником фактора форми, який наближений до 1,0 (табл. 1). Звертає на себе увагу той факт, що гіпертрофія та гіперплазія мітохондрій у симпластах литкового м’яза після фізичного навантаження супроводжується значним зниженням числа гранул глікогену, тобто у цих тварин відбувається зміна гліколітичного шляху метаболізму на окислювально-відновний. Про функціонування мітохондрій у напруженому режимі свідчить і наявність у них великої кількості розташованих, в основному паралельно, крист.
Мітохондрії мають свої особливості як за кількісним представництвом, так і ультраструктурою. В об'ємі симпласту, що дорівнює 1,0 мкм3, міститься (39,62±5,78) х 10-2 мітохондрій при середній площі зрізу (30,18±2,01) х 10-2 мкм2. Слід відзначити, що, хоча за середнім показником кількісна щільність мітохондрій у цих щурів статистично не відрізняється від інтактних (табл. 1), внаслідок великої помилки через значний розкид показників у експериментальних тварин, розподіл ділянок з різним числом цих органел суттєво різниться між собою. В ІІ підгрупі щурів після фізичного навантаження, як і у контрольних тварин, більша частина симпласту містить у 1 мкм3 до 202 мітохондрій.

Рис. 3. М’язові волокна у камбалоподібному м’язі щурів після довготривалого фізичного навантаження. Мітохондрії з концентричним розташуванням крист. Електронномікроскопічне фото. Зб. 48 000.
Одночасно в ультраструктурі м’яза з’являються ділянки (~ 20%), що переповнені мітохондріями. Такі ділянки у тварин І підгрупи, де виражені компенсаторно-пристосувальні процеси, становлять > 50%, через що величина кількісної щільності суттєво (більш ніж удвічі) і достовірно перевищує середній показник у ІІ підгрупі. Відмінності мітохондрій за площею зрізу в І та ІІ пі-Оригінальні дослідження в групах також є значущими, але не такими виразними, як кількість цих субклітинних органел. Розподіл мітохондрій за площею в інтактних щурів має несиметричний характер; в І підгрупі основної групи він найбільш близький до нормального, властивого інтактним тваринам, тоді як у ІІ підгрупі цей показник зазнає незначного зсуву вліво за відсутності одного проміжного класу.
Окрім кількісних показників, мітохондрії у порівнювальних групах відрізняються також за ультраструктурою. Внаслідок інвагінацій на своїй поверхні вони часто набувають неправильної форми, що підтверджується і результатами морфометричного аналізу. Величина фактора форми зменшується до 0,75±0,01, що достовірно менше, ніж у контролі та І підгрупі (табл. 1). Більшість мітохондрій не мають чітко структурованої зовнішньої мембрани; кристи, як правило, фрагментовані, вакуолізовані, число їх незначне; матрикс гомогенізований. Ще однією особливістю таких мітохондрій є накопичення в них гранул кальцію (рис. 2).
Кількість гранул глікогену всюди в симпластах є незначною, що суттєво відрізняє їх від інтактних тварин і на тлі поширеності деструктивнодистрофічних змін мітохондрій може відображати стадію декомпенсації в енергетичному забезпеченні волокон у литковому м’язі деяких щурів при довготривалому фізичному навантаженні.
Мітохондрії набувають неправильної форми, мають електронно-прозорий матрикс та паралельно розташовані кристи. Іноді спостерігаються органели з центричним розташуванням крист (рис. 3), що характерно при напружених процесах. Морфометричний аналіз показав, що фізичне навантаження викликає збільшення розмірів мітохондрій камбалоподібного м’яза. Середня площа цих органел більш ніж удвічі перевищує цей же показник у інтактних тварин. Причому, як і у контролі, мітохондрії у підсарколемній зоні більші за площею зрізу, ніж ті, що розташовані міжфібрилярно (табл. 2).
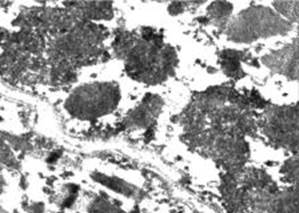
Рис. 2. М’язові волокна у литковому м’язі щурів групиІІ після довготривалого фізичного навантаження. Мітохондрії із гомогенізованим матриксом помірної електронної щільності, залишками крист та гранулами кальцію. Електронномікроскопічне фото. Зб. 48 000.
Таке збільшення відбувається внаслідок зниження числа дрібних мітохондрій і появі крупних органел, які не спостерігаються у контролі. Фактор форми мітохондрій віддаляється від 1, тобто вони втрачають кулясту або овальну форму. Більш виразних змін зазнають органели у міжфібрилярних ділянках. Якісний аналіз свідчить, що такі зміни відбуваються через формування на поверхні мітохондрій значних випинів. Кількісна щільність мітохондрій за середніми показниками значно не відрізняється від контролю в обох зонах, але при фізичному навантаженні похибка середньої величини є дуже великою. Це є результатом нерівномірного розподілу мітохондрій у різних ділянках симпласту. При фізичному навантаженні і підсарколемно, і міжфібрилярно переважають ділянки з найменшим числом цих органел (до 20×10-2 в мкм3). Водночас з’являються ділянки, де їх кількість значна (60−80×10-2/мкм3), тоді як у інтактних тварин переважають ділянки з середнім числом (20−40×10-2/мкм3) мітохондрій. Хоч загальна кількість мітохондрій у м’язових волокнах камбалоподібного м’яза не збільшується, за рахунок їх значної гіпертрофії значно (майже в 4 рази) збільшується їх об'ємна щільність. Як і у контролі, більший об'єм мітохондрії займають у підсарколемній зоні симпластів (табл. 2).
Тобто, литковий та камбалоподібний м’язи у різних експериментальних тварин за однакового тренування по-різному реагують на довготривале фізичне навантаження. У частини тварин воно викликає розвиток виразних адаптаційних процесів без суттєвих пошкоджень ультраструктур, тоді як у інших переважають деструктивно-дистрофічні процеси у м’язових волокнах. Це, можливо, пов’язано з особливостями будови литкового м’яза, який належить до так званого «білого» типу, та камбалаподібного м’яза, який належить до так званого «червоного» типу [7]. Відомо, що м’язи складаються з різних типів волокон: 1 тип — це м’язові волокна, які скорочуються повільно і мають дуже високу опірність до стомлення. До типу 2 належать волокна, які скорочуються швидше, ніж волокна 1 -го типу, але тоді ж і швидше втомлюються. Швидкі м’язові волокна типу, А виробляють більше потужності, ніж повільні, але відстають за витривалістю від волокон типу B. Цей тип В ще називають «проміжним», оскільки йому притаманні властивості як швидких, так і повільних волокон [8]. Залежно від їхньої специфічної комбінації литковий та камбалоподібний м’язи у різних тварин будуть по-різному реагувати на довготривале фізичне навантаження, що і спостерігається в нашому дослідженні, а також підлягають модерації залежно від побудови тренувального процесу, раціону, фармзабезпечення тощо. Специфічні фізичні навантаження значної тривалості у процесі адаптації можуть викликати експресію визначених генів, зокрема mRNA інтерлейкіну 6 (IL-6), рецептора IL-6, інсуліноподібного фактора росту, фосфофруктокінази та транспорту глюкози, кінцевим наслідком чого стає покращення резистентності організму до негативних зовнішніх чинників [9].
Таблиця 2Морфометричні показники мітохондрій у периферійних (І) та центральних (ІІ) ділянках симпластів камбалоподібного м’яза щурів контрольної та основної груп.
Групи. | Об'ємна щільність МТ, %. | Кількісна щільність МТ, 10-2/мкм3 | Площа зрізу МТ, 10-2 мкм2 | Фактор форми. | |
К-I. | 4,73+1,83. | 29,85+1,03. | 18,01+1,61. | 0,80+0,1. | |
К-II. | 2,72+0,93. | 24,73+1,54. | 13,97+1,16. | 0,80+0,1. | |
ФН-І. | 17,49+7,07. | 29,44+9,27. | 42,49+2,54. | 0,78+0,01. | |
ФН-ІІ. | 9,10+4,24. | 22,57+11,75. | 33,76+4,31. | 0,69+0,02. | |
Висновок
Такий розподіл змін ультраструктури мітохондрій литкового та камбалоподібного м’язів у різних експериментальних тварин, що перебували під впливом фізичного навантаження однакової тривалості та інтенсивності, свідчить про індивідуальні пристосувальні механізми різної виразності та сили до тривалого навантаження та прояснює механізми адаптаційних перебудов на рівні ультраструктури м’язового волокна та показників, що відображають напруженість енергогенеруючих процесів у скелетних м’язах.
Перспективи подальших досліджень. Отримані результати свідчать про необхідність подальшого вивчення змін у м’язовій системі при фізичних навантаженнях, шляхів корекції виявлених змін, а також при застосуванні різноманітних факторів, які позитивно впливають на фізичну працездатність, що допоможе створити підґрунтя для адекватного використання засобів корекції біоенергетичних процесів в організмі.