Мінеральний склад кореневищ перстача прямостоячого (potentilla erecta L.)

Селен не є необхідним елементом для рослин, але перебуває у всіх його частинах у вигляді селеновмісних амінокислот і частково у вигляді селеніті селенат-іонів. Селен у рослинах захищає їх від мікробних патогенів та поїдання тваринами, однак він не є суттєвим для вищих рослин. При підвищених рівнях Se є токсичним для більшості рослин, через неспецифічне включення Se в сполуки сірки, оскільки вони… Читати ще >
Мінеральний склад кореневищ перстача прямостоячого (potentilla erecta L.) (реферат, курсова, диплом, контрольна)
Пріоритетні дослідження лікарських рослин традиційно здійснюють стосовно їхнього біохімічного складу та дії біологічно активних речовин.
Останнім часом спостерігається зростання інтересу до вивчення їхнього макрота мікроелементного складу, оскільки ці елементи містяться в каталітичних центрах молекул ферментів і здатні регулювати їхню активність, тим самим істотно впливати на біохімічні процеси в клітинах. Низка елементів присутня майже в усіх рослинах як мікроелементи і має важливе значення для метаболізму рослин у невеликих кількостях (Mn, Fе, Cu, Mo, Zn, Ni). У високих концентраціях ці метали є токсичними. Такі елементи як Na, Co, S, Mg тощо стимулюють ріст і мають важливе значення тільки для певних видів рослин. Тому якісний і кількісний елементний склад рослин є видоспецифічною ознакою.
Мінеральні речовини, які вибірково накопичують рослини, можуть впливати на їхню фармакологічну дію: підвищувати або знижувати їхню всмоктуваність, резорбтивні властивості, бути синергістами або антагоністами, а також зменшувати чи посилювати токсичну дію. Разом з тим деякі хімічні елементи можуть модулювати вироблення вторинних метаболітів (алкалоїдів, терпеноїдів, фенольних сполук, сапонінів, полісахаридів, флавоноїдів) залежно від виду рослин, а також від діючої концентрації цих елементів [1, 2]. Так, показано зростання кількості вторинних метаболітів під впливом Ni, Ag, Fe, Co у Cynoglossum furcatum Wallich, Elsholtzia blanda Bentham, Lycopodium cernuum Linnaeus, Potentilla fulgens Wallich ex Hooker, Swertia macrosperma C. B. Clarke, Thalictrum foliolosum DC, Valeriana jatamansi Jones [3, 2, 4].
У багатьох біохімічних процесах у рослинному організмі беруть участь флавопротеїнові ферменти. В активації цих ферментів беруть участь Mn, Fe, Cu, Mo. Для вивчення синтезу біологічно активних речовин найбільший інтерес попередньо являють Mn, Mo, Cu, Co, Ni, Sr, V і Cr. Вважають, що лікарські рослини, які продукують дубильні речовини, вибірково накопичують Mn, Cu, Cr. Високі концентрації Mn забезпечують синтез аскорбінової кислоти і танідів, кількість яких корелює з накопиченням Mn у рослинах. Окрім того, високі концентрації низки макрота мікроелементів у лікарських рослинах можуть зумовити їхню токсичність [5]. Деякі мікроелементи, такі як миш’як, кадмій, свинець і ртуть, розглядають як шкідливі для людського організму, тому важливо, щоб вони були відсутні у лікарській рослинній сировині чи були наявні в допустимих концентраціях.
Вибіркова властивість рослин до високої концентрації хімічних елементів є видоспецифічною ознакою. За здатністю накопичувати певні елементи відрізняють види рослин, які є концентраторами Zn, Co, Mn, Mg тощо [5]. Доступність для рослин хімічних елементів ґрунту залежить як від біології видів (зокрема вибірковості поглинання кореневими системами і метаболічних процесів у тканинах), так і від хімічного складу оселищ. Тому вміст певних елементів у різних видах рослин з одного екотопу відрізняється, що властиве також рослинам одного виду, зібраним із різних локалітетів [4, 6, 1]. Останнє підтверджується нашими дослідженнями.
Potentilla erecta (перстач прямостоячий, калган) належить до найбільш досліджених лікарських рослин роду Potentilla, для надземних і підземних частин якого встановлено в цілому 68 біологічно активних речовин, зокрема 43 хімічні сполуки ідентифіковано в кореневищах Potentilla erecta [7]. Найбільшу групу становлять таніни, загальний вміст яких варіює у межах 17−22%. Вони включають у себе головним чином конденсовані дубильні речовини і гідролізовані дубильні речовини. Важливими компонентами також є катехіни та тритерпеноїди. У незначних концентраціях присутні флавоноїди, зокрема органічні, і гідроксикоричні кислоти. Природне поєднання цих сполук спричинює виражену антимікробну дію відносно грампозитивних бактерій, деяких грибів та грамнегативних бактерій [7, 8]. Досліджено також цитотоксичну активність екстрактів Potentilla erecta проти вірусів герпесу, грипу, коров’ячої віспи, ротавірусу. Такі цілющі властивості діючих речовин Potentilla erecta зумовили широке використання її офіційною медициною багатьох країн.
Монографії на сировину Potentilla erecta включено до Європейської, Австрійської, Чеської фармакопей, Британської трав’яної фармакопеї, Державної фармакопеї України. Використання її сировини для медичних чи фармацевтичних цілей має офіційний статус також у Литві, Латвії, Польщі, Бельгії та Іспанії.
Широкий спектр протизапальної дії Potentilla erecta пояснюється в основному наявністю флавоноїдів, і може бути пов’язаний з їхньою антиоксидантною активністю, яка посилюється іншими органічними і неорганічними (макроі мікроелементами — Fe, Mg, Mn, Zn та ін.) сполуками. Якщо біологічно активні речовини Potentilla erecta та їхня дія досліджені досить детально, то елементний склад сировини цього виду рослин вивчений мало.
Метою нашого дослідження було вивчити елементний склад рослинної сировини Potentilla erecta у межах її сировинної зони з географічно віддалених територій України — українського Полісся та українських Карпат.
Об'єктом дослідження були кореневища Potentilla erecta з різних природних зон України — українських Карпат (зразок 1) та українського Полісся (зразок 2). Зразок 1 зібрано на полонині поблизу с. Яблуниця на висоті близько 1 000 м н. р. м.; зразок 2 — у сосновому лісі біля с. Миколаївка Макарівського району Київської обл. Значення рН водної витяжки ґрунту з місця збору зразка 1 становило 4,2, зразка 2 — 5,7. Зібрані кореневища підсушували на відкритому повітрі, зважували. Вологість зразків не перевищувала 12−15%. Для аналізу брали 0,1 г зразка, додавали 2,0 мл концентрованої HNO3 (Merck) і піддавали мінералізації закритим способом у мікрохвильовій печі MWS-2 фірми Berghof. Отриманий прозорий мінералізат розчиняли в деіонізірованій воді (18Q) до об'єму 10 мл. Отриманий мінералізат аналізували методом атомно-емісійної спектрометрії з індуктивно-зв'язаною плазмою (Optima 210 DV фірми Perkin Elmer, США). Робота спектрометра управляється і контролюється програмним забезпеченням WinLab32 в операційній системі Windows XP prof. Одержані результати обробляли приладом автоматично і виводили на монітор у необхідному форматі у вигляді протоколу. Похибка вимірювання становить близько 2%.
У досліджених зразках кореневищ Potentilla erecta виявлено 29 елементів, кількісні показники яких характеризуються значною варіабельністю (таблиця). Рівні розподілу аналізованих елементів у кореневищах загалом можна розмістити у порядку зменшення таким чином: Ca > S > Mg > K > Sn > I > P > Ba > Al > Zn > Mn > Fe > Sr > Si > Ag > Cr > Ni > Cu > Pb > V > Se > Ti > Cd > Ge > Na > Mo > Co > As > Bi.
Т, а б л и ц я Вміст хімічних елементів у кореневищі Potentilla erecta (мг/кг сухої маси).
Елемент. | Зразок 1. | Зразок 2. | |
Ca. | 3 117,51 ± 623,5. | 2 305,61 ± 461,12. | |
S. | 652,93 ± 130,59. | 938,04 ± 187,60. | |
Mg. | 672,79 ± 134,56. | 637,34 ± 127,47. | |
K. | 461,40 ± 92,28. | 274,10 ± 54,82. | |
Sn. | 290 ± 30,46. | 310,49 ± 62,09. | |
I. | 381,70 ± 76,34. | 147,90 ± 29,58. | |
P. | 172,05 ± 34,41. | 200,58 ± 40,12. | |
Ba. | 270,69 ± 54,14. | 59,54 ± 11,01. | |
Al. | 124,45 ± 24,89. | 21,97 ± 4,39. | |
Zn. | 79,13 ± 15,83. | 12,21 ± 2,44. | |
Mn. | 51,78 ± 10,36. | 10,64 ± 2,13. | |
Fe. | 32,34 ± 6,47. | 3,27 ± 0,65. | |
Sr. | 19,17 ± 3,83. | 12,25 ± 2,45. | |
Si. | 12,01 ± 2,40. | 4,10 ± 0,82. | |
Ag. | 2,88 ± 0,58. | 6,46 ± 1,29. | |
Cr. | 2,65 ± 0,53. | 3,05 ± 0,70. | |
Ni. | 2,73 ± 0,55. | 1,09 ± 0,22. | |
Cu. | 1,75 ± 0,35. | 1,48 ± 0,30. | |
Pb. | 2,06 ± 0,41. | 0,36 ± 0,07. | |
V. | 0,48 ± 0,10. | 0,52 ± 0,10. | |
Se. | 0,67 ± 0,13. | 0,08 ± 0,02. | |
Ti. | 0,11 ± 0,02. | 0,60 ± 0,12. | |
Cd. | 0,59 ± 0,11. | 0,10 ± 0,01. | |
Ge. | 0,48 ± 0,09. | 0,14 ± 0,03. | |
Na. | 0,42 ± 0,08. | 0,14 ± 0,03. | |
Mo. | 0,10 ± 0,02. | 0,34 ± 0,07. | |
Co. | 0,18 ± 0,04. | 0,04 ± 0,008. | |
As. | 0,003. | 0,003. | |
Bi. | 0,002. | 0,002. | |
Результати аналізу свідчать, що серед макроелементів перстач прямостоячий найбільше накопичує кальцій, сірку і магній. Причому, вміст магнію майже не відрізнявся в обох зразках, тоді як у зразку 2 виявлено кальцію майже на чверть менше, а сірки на третину більше, ніж у зразка 1 (рисунок, а). Очевидно, що такі відхилення у накопиченні аналізованих елементів пов’язані з відмінностями кліматичних умов оселищ та властивостями грунту, що може впливати також на зміну фізіологічних процесів рослин, зокрема вторинний метаболізм [1, 2, 4].
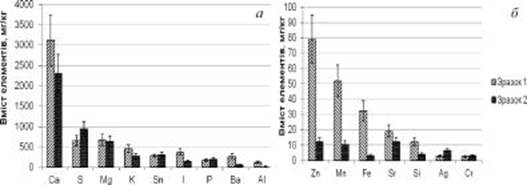
Cu Ni Pb V Se Ті Cd Ge Na Mo Со Рис. Вміст макрота мікроелементів у кореневищах Potentilla erecta
K, Ca і Mg належать до макроелементів вищих рослин через велику роль цих елементів для росту рослин. У рослинній клітині Са присутній в основному в клітинній стінці і в вакуолях, де він бере участь в стабілізації клітинної стінки і секреторних процесах. Са досить нерухомий в флоемі, тому він накопичується в старих частинах рослин. Деякі рослини природно багаті кальцієм, як кропива дводомна. У маку (Papaver somniferum) концентрація Са в насінні та листі вище, ніж концентрація K [9].
Вміст кальцію в кореневищах карпатських рослин калгану становить 3 117,51 ± 623 мг/кг, а у поліському зразку — 5 2305,61 ± 461,12 мг/кг Показники кальцію є найвищими серед аналізованих елементів кореневищ Potentilla erecta, вони майже втричі перевищують вміст сірки та магнію. Зазначимо, що вибіркова властивість накопичення у великій кількості Са, S, Mg та K притаманна також кореневищам Potentilla alba.
Відомо, що Са належить до основних складових хімічних елементів також для надземної частини калгану. За різними даними, у кількісному вираженні він займає 1-е місце [6], або 2−3-є після К та Na [1, 4]. Визначено, що високий вміст кальцію притаманний рослинам, які мають кровоспинну дію.
Сірка є одним з найважливіших макроелементів для рослин, і її різні форми необхідні в проміжному метаболізмі рослини. Потреба рослин у сірці залежить від рівня азотного живлення. Чим вище потреба азоту, тим більше сірки вони споживають. Це може бути однією з основних причин відмінності показників її кількісного вмісту в аналізованих зразках. У житті рослин сірка відіграє важливу фізіологічну роль, входячи до складу багатьох речовин, необхідних для життєдіяльності рослин і важливу роль у фотосинтезі та вторинному метаболізмі сірковмісних сполук переважно фенольної природи, які прямо або побічно пов’язані зі захистом рослин від патогенних мікроорганізмів.
Разом із магнієм сірка є кофактором багатьох ферментів, які беруть участь в енергетичному обміні, синтезі білків тощо. Дефіцит сірки в рослинах впливає на метаболізм азоту і, у зв’язку з цим, на розвиток всієї рослини. Дослідженню фармацевтичної цінності сірковмісних продуктів рослинного походження надається останнім часом значна увага. Встановлено, що накопичення цього елементу притаманне переважно представникам родин цибулевих, хрестоцвітих, бобових, макових [10]. Даних щодо вмісту сірки у різних органах роду Potentilla обмаль, але відомо, що сірка та магній тісно корелюють зі зміною фенольних метаболітів у рослинах [11].
Магній входить до складу хлорофілу, бере участь в активації багатьох ферментів. Діапазон вмісту Mg у зелених тканинах становить від 0,15 до 0,35% в розрахунку на суху речовину, тому вміст Mg більше в листі, ніж в інших органах рослин [9]. Результати наших досліджень свідчать, що P. erecta притаманна властивість накопичувати в кореневищах велику кількість магнію у різних екологічних умовах зростання (таблиця).
Показники вмісту К, І, Ba i Al у карпатському зразку значно перевищують такі у поліському (рисунок, а), тоді як вміст Sn та P дещо вищий у останньому. Такі відмінності, очевидно, зумовлені різними екологічними умовами оселищ, зокрема ступенем екологічного забруднення середовища зростання рослин, оскільки зразок № 2 зібраний у лісовому масиві на відстані близько 35 км від Києва, а зразок № 1 — в екологічно чистому регіоні.
Калій зазвичай знаходиться в високих концентраціях в тканинах рослин. Він бере участь у процесах первинного та вторинного метаболізму, включаючи активацію ферментів, синтез білка, фотосинтез, транспорт органічних сполук тощо. Для оптимального росту концентрація K становить 2−5% від сухої маси рослин.
Немає ніяких доказів того, що олово є істотним або корисним для рослин, але рослини можуть легко вбирати Sn із ґрунту, при тому велика частина поглиненого Sn залишається в коренях. Відомо, що на мінералізованих забруднених ґрунтах рослини здатні накопичувати велику кількість олова і можуть бути токсичними [5]. Здатність накопичувати калганом олово має ознаки стабільності, оскільки кількісні показники його вмісту майже не відрізняються у обох зразках (таблиця).
Барій не є істотним компонентом рослинних тканин, але часто присутній у рослинних тканинах разом із К. На кислих грунтах Ва краще поглинається рослинами. Він є антагоністом кальція, магнія та сірки [5, 9]. Кількісні показники вмісту Ва у карпатському зразку майже у п’ять разів перевищують такі у поліському зразку. Такий широкий діапазон накопичення барію різними органами лікарських рослин притаманний також іншим видам. Дані літератури щодо накопичення рослинами барію та йоду неоднозначні, хоча є дані про позитивний вплив йоду на накопичення фенольних сполук у рослинах [12]. В аналізованих зразках вміст барію суттєво відрізняється (рисунок, а), що може свідчити про залежність накопичення цього елементу від умов зовнішнього середовища.
Фосфор є важливим макроелементом рослин і становить близько 0,2% від сухої маси рослини. Він є компонентом нуклеїнових кислот, фосфоліпідів і АТФ, а також бере участь у контролі ключових ферментативних реакцій і в регуляції метаболічних процесів. Вміст Р в аналізованих нами зразках невеликий (таблиця) і кількісно майже не відрізняється.
Вміст алюмінію в рослинах сильно варіює залежно від ґрунтових умов [9]. Фізіологічна функція Al в рослинах не з’ясована чітко, хоча є деякі докази того, що низький вміст Al може сприятливо впливати на ріст рослин, зокрема активувати деякі ферменти [5]. Токсичність Al для рослин більш виражена на кислих грунтах, що є основною перешкодою для вирощування сільськогосподарських культур. Коли рН ґрунту нижче 5, алюміній вивільняється в ґрунт і входить у клітини кореневого чохлика, при цьому спричинює пригнічення подовження і ділення клітин, що призводить до кореневої низькорослості, супроводжуваної зменшенням поглинання води і поживних речовин [5]. Дуже високі показники накопичення Al виявлені в деяких лікарських рослинах. Наприклад, у шавлії вони сягають рівня 1 722 мг/кг, у коріандрі - 1 446 мг/кг [13]. Рівень накопичення Al у наших зразках P erecta незначний і становить 21,97−124,45 мг/кг (таблиця).
Серед інших елементів у аналізованих кореневищах Potentilla erecta найбільше виявлено цинку, марганцю і заліза (рисунок, б). Водночас для цих елементів наявна найбільша диспропорція їхніх кількісних показників у аналізованих зразках. У поліському зразку Fe менше, ніж у карпатському у 10 разів, Zn — у 6, а Mn — у 5 разів (таблиця). Очевидно, такий широкий діапазон накопичення цих елементів зумовлений екологічними умовами оселищ.
Відомо, що надземна частина P erecta серед цих елементів найбільше накопичує заліза, кількість якого в надземній частині рослин перевищує вміст Zn і Mn у десятки разів [4]. В інших дослідженнях [6], вміст Fe перевищує вміст Zn у надземній частині рослин Potentilla erecta у 15 разів. Такі дані свідчать про можливі значні варіації кількісного вмісту цих елементів у різних умовах зростання рослин.
Значення цинку для росту рослин тісно пов’язано з його участю в азотному обміні. Його фізіологічна роль у рослинах багатогранна. Цинк відіграє важливу роль в окисно-відновних процесах, що відбуваються у рослинному організмі, він є складовою частиною ферментів, безпосередньо бере участь у синтезі хлорофілу, впливає на вуглеводний обмін у рослинах і сприяє синтезу вітамінів. Постійний компонент ферментів для синтезу білка і вироблення енергії, відіграє важливу роль у розвитку насіння. Синтез вторинних метаболітів, таких як поліфенольні сполуки, зокрема танін, є одним із найважливіших показників активності окисно-відновних процесів та ферментативних реакцій за участю цинку. Присутність цинку сприяє вуглеводневому обміну, а також підвищенню вмісту таніну. Високі його концентрації у рослинах характерні переважно для екологічно забруднених регіонів. Деякі види роду Potentilla вважають гіперакумуляторами цинку.
Марганець необхідний для метаболізму і розвитку рослин. Він відіграє важливу роль в активуванні ферментів, що впливають на утворення ароматичних амінокислот, лігнінів, флавоноїдів і фітогормонів. Марганець бере участь не лише у фотосинтезі, а й у синтезі вітаміну С. Застосування мікродобрива з Mn збільшує вміст катехінів і флавоноїдів. Роль марганцю в обміні речовин у рослин подібна до функцій магнію і заліза.
Залізо бере участь у процесі фотосинтезу, мітохондріального дихання, засвоєння азоту, біосинтезі гормонів, виробництва та очищення активних форм кисню, захисту від патогенних впливів навколишнього середовища. До 80% від клітинного заліза знаходиться в хлоропластах, що відповідає його основній функції в процесі фотосинтезу. Фенольні сполуки накопичуються в тканинах, багатих на залізо і цей факт може бути пов’язаний з механізмом мобілізації Fe в рослинах, оскільки феноли можуть утворювати комплекс із залізом. Серед видів роду Potentilla, перстач прямостоячий має здатність накопичувати багато заліза і в надземних органах [4, 6].
Роль кремнію, стронцію, срібла та хрому у фізіологічних процесах рослин детально не з’ясовано. Припускається, що іони срібла захищають рослини від грибкових, вірусних і мікробних захворювань [10]. Стронцій бере участь у синтезі певних груп БАР, у малих дозах він виконує функції, притаманні кальцію [5]. Кремній має суттєвий вплив на поглинання та використання рослинами інших елементів мінерального живлення. Є дані про позитивний вплив кремнію на поглинання рослинами азоту. Під його впливом зростає асиміляція рослинами калію, кальцію і магнію. У присутності кремнію рослини ефективніше використовують бор і можуть легше переносити надлишок марганцю, алюмінію і заліза. Позитивний вплив Cr на фотосинтез дає змогу припустити вплив цього елемента на продукування біологічно активних речовин. У флавоноїдовмісних рослин найбільша кількість Cr припадає на органи, багаті флавоноїдами.
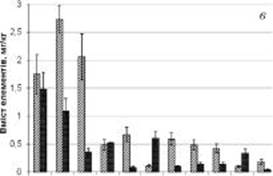
При аналізі вмісту мікрота макроелементів у кореневищах Potentilla erecta виявлено, що вони акумулюють незначно також Ni, Cu та Pb (рисунок, в). Високий вміст Ni відзначено в квітках флавоноїдовмісних рослин. P. erecta належить до видів лікарських рослин, для яких характерна вибіркова властивість накопичення міді. Поєднання дубильних речовин із підвищеним вмістом Cu встановлено у багатьох лікарських рослин, які використовують для лікування численних захворювань, пов’язаних із запаленням різних тканин тіла людини. Мідь у рослинах тісно пов’язана з фотосинтезом та виробництвом фенольних сполук. Вважається, що цей метал є важливим активатором ферменту для синтезу фенольних кислот у тканинах рослин. Присутність у невеликій кількості свинцю у рослинах позитивно впливає на фотосинтез [5], однак надмірна його кількість, що притаманно екологічно забрудненим регіонам, викликає пригнічення росту та розвитку рослин [10] і збільшення токсичності лікарської рослинної сировини. Послідовність зменшення вмісту цих елементів (Ni > Cu > Pb) в аналізованих кореневищах калгану подібна до такої у надземній частині деревію звичайного [14].
Вміст інших мікроелементів у досліджуваних кореневищах Potentilla erecta мінімальний і варіює у межах 1 мг/кг (таблиця). Дисбаланс кількісних показників цих мікроелементів ще різкіший (рисунок, в). У карпатському зразку майже у девять разів більше селену, у шість — кадмію, у три — германію та натрію (таблиця). Разом з тим у поліському зразку більше, ніж у п’ять разів, титану та у три — молібдену, а ванадію — майже однакова кількість. Така різниця у показниках може свідчити про визначальну роль середовища у накопиченні слідових хімічних елементів цим видом рослин.
Більшість аналізованих мікроелементів у мінімальній кількості позитивно впливають на метаболізм рослинних клітин. Так, ванадій впливає на фотосинтез, азотний і водний обмін. Його було ідентифіковано у багатьох протиракових лікарських рослинах. Разом з тим, ванадій, селен, титан, свинець та кадмій — високотоксичні елементи. Селен, як і залізо та цинк, є важливим компонентом ферментів, які беруть участь у виробленні первинних та вторинних метаболітів.
Селен не є необхідним елементом для рослин, але перебуває у всіх його частинах у вигляді селеновмісних амінокислот і частково у вигляді селеніті селенат-іонів. Селен у рослинах захищає їх від мікробних патогенів та поїдання тваринами, однак він не є суттєвим для вищих рослин. При підвищених рівнях Se є токсичним для більшості рослин, через неспецифічне включення Se в сполуки сірки, оскільки вони хімічно подібні. Гіперакумуляторами селену є деякі представники родин Brassicaceae, Fabaceae та Asteraceae. перстач potentilla рослинний сировина Титан спричинює стимулювальний вплив на метаболізм рослин шляхом підвищення активності деяких ферментів і збільшення концентрації заліза у хлоропластах. Він, як і нікель, належить до елементів із низькою інтенсивністю переходу з ґрунту в рослини. Кадмій (Cd) належить до так званих важких металів, які входять у групу особливо небезпечних речовин. Він легко всмоктується з ґрунту через кореневу систему, а також з атмосфери. Cd не належить до необхідних для життєдіяльності рослин елементів і здатний порушувати роботу ферментів. Будучи токсичним за відносно низьких концентрацій, він легко поглинається і поширюється у тканинах рослин. Однак рослини, акумулятори сірки, мають природний захист клітин від токсичного впливу Cd. Кадмій потрапляє в ґрунт за допомогою різних антропогенних забруднень (зокрема у результаті застосування фосфорних добрив, гною, викиду стічних вод, транспорту тощо). Існує непряма лінійна залежність між рН ґрунту і Cd-біодоступністю, тобто зі зменшенням рН ґрунту поглинання рослинами металу збільшується. Однак присутність цих елементів у мікродозах в аналізованій сировині не може нести загрози здоров’ю людини.
Молібден за своїм практичним значенням для рослин займає одне з перших місць серед інших мікроелементів. Під впливом молібдену в рослинах збільшується вміст хлорофілу і підвищується інтенсивність фотосинтезу, збільшується вміст вуглеводів, каротину і аскорбінової кислоти, підвищується вміст білкових речовин [1]. Потреба рослин у молібденових добривах зазвичай зростає на кислих ґрунтах, що мають рН нижче 5,2 [3]. Зазначимо, що вміст Мо у поліському зразку втричі перевищував показники карпатського зразка (рисунок, в).
Кобальт є важливим компонентом низки ферментів і коферментів. Він впливає на ріст і метаболізм рослин залежно від концентрації і наявності кобальту в ризосфері і ґрунті. Наприклад, високий рівень Со спричинює дефіцит заліза в рослинах і пригнічує поглинання Cd корінням. Він також взаємодіє синергічно з Zn, Cr і Sn. Сприятливий вплив кобальту — уповільнення старіння листя, підвищення стійкості до посухи в насінні, регуляція накопичення алкалоїдів у лікарських рослинах, а також пригнічення біосинтезу етилену. У нашому експерименті вміст Со у кореневищах калгану був мінімальним, особливо у поліському зразку (таблиця).
У кореневищах калгану також виявлено сліди миш’яку та вісмуту.
Висновок
Для визначення ефективності рослин під час лікування різних захворювань важливо зрозуміти їхню фармакологічну дію з урахуванням інформації про здатність концентрувати у сировинних органах певні хімічні елементи. Встановлено, що Potentilla erecta є концентратором Са, S, Mg. У кількості більше 100 мг/кг ідентифіковано K, Sn, I, P, Ba та Al. Вміст інших елементів незначний. Кількісні дані деяких металів відрізняються у різних аналізованих зразках. Основною причиною цього, ймовірно, є розбіжність екологічних умов оселищ у місцях збору сировини. Одержані експериментальні дані можуть бути використані для прогнозування і планування фармакогностичних досліджень Potentilla erecta та розроблення методики контролю якості та стандартизації на рослинну сировину цієї лікарської рослини.
Література
Broadley M. R., Bowen H. C., Cotterill H. L. et al. Phylogenetic variation in the Shoot mineral concentration of angiosperms // J. Experimental Botany. — 2004. — V. 55, N 396. — P. 321−336. Downloaded from http://jxb.oxfordjournals.org/by guest on November 17, 2016.
Hassan A. Effects of Mineral Nutrients on Physiological and Biochemical Processes Related to Secondary Metabolites Production in Medicinal Herbs. Medicinal and Aromatic Plant Science and Biotechnology 6 (Special Issue 1), 2012. — P. 105−110. http://dx.doi. org/10.1590/S0102−695X2011005000177.
Anal J. M. H., Chase Р. Trace elements analysis in some medicinal plants using graphite furnace — Atomic absorption spectroscopy Environ. Eng. Res. — 2016 http://dx.doi. org/10.4491/eer.2016.007.
РупасоваЖ. А., Рудаковская Р. Н., Матюшевская Е. Н. и др. Сравнительная оценка видов рода Potentilla L. по минеральному составу надземных органов в условиях Беларуси // Известия Нац. академии наук Беларуси. Сер. Биологич. наук. — 2002. — № 3. — С. 5−9.
Kabata-Pendias A., Pendias H. Trace elements in soils and plants / CRC Press LLC.2001. — 403р.
Остапко И. Н. Сравнительный анализ содержания элементов полезных растений из коллекции Донецкого ботанического сада НАН Украины // Промышленная ботаника. — 2003. — Вып. 3. — С. 87−90.
TomczykM., Bazylko A., Staszewska A. Determination of polyphenolics in extracts of Potentilla species by high-performance thin-layer chromatography photodensitometry method. — 2010. — V. 21, Issue 2. — P. 174−179.
Synowiec A., Gniewosz M., Bqczek K., Przybyi J. L. Antimicrobial effect of an aqueous extract of Potentilla erecta rhizome // Herba Polonica. — 2014. — V. 60, N 2.
Chizzola R. Metallic Mineral Elements and Heavy Metals in Medicinal Plants Medicinal and Aromatic Plant Science and Biotechnology 6 (Special Issue 1). — 2012. — Р. 39−53.
Коновалова О. Ю., Мітченко Ф. А., Шураєва Т. К., Джан Т. В. Мінеральні елементи лікарських рослин та їх роль у життєдіяльності людини. — К.: Видавничополіграфічний центр «Київський університет», 2012. — 192 с.
Sampaio B. L., Bara M. T. F., Ferri P H. et al. Influence of environmental factors on the concentration of phenolic compounds in leaves of Lafoensia pacari // Rev. bras. farmacogn. — 2011. — V. 21, N 6. — Epub Sep 30, 2011.
Blasco B., Leyva R., Romero L., Ruiz J. M. Iodine effects on phenolic metabolism in lettuce plants under salt stress // J. Agric. Food Chem. — 2013. — V. 20, N 61 (11). — P. 2591−2596. doi: 10.1021/jf303917n. Epub 2013 Mar 8.
Zengin M., Ozcan M. M., Cetin U., Gezgin S. Mineral contents of some aromatic plants, their growth soils and infusions // J. Science of Food and Agriculture. — 2008. — V. 88. — Р 581−589.
Семенова В. В. Видовая специфика накопления тяжелых металлов лекарственными растениями // Всерос. журн. научных публикаций. — 2011. — № 1 (2). — С. 17−18.